

Review
doi: 10.1016/j.neuron.2009.04.008.
Rapid neocortical dynamics: cellular and network mechanisms
Affiliations
- PMID: 19409263
- PMCID: PMC3132648
- DOI: 10.1016/j.neuron.2009.04.008
Item in Clipboard
Review
Rapid neocortical dynamics: cellular and network mechanisms
Neuron.
.
Abstract
The highly interconnected local and large-scale networks of the neocortical sheet rapidly and dynamically modulate their functional connectivity according to behavioral demands. This basic operating principle of the neocortex is mediated by the continuously changing flow of excitatory and inhibitory synaptic barrages that not only control participation of neurons in networks but also define the networks themselves. The rapid control of neuronal responsiveness via synaptic bombardment is a fundamental property of cortical dynamics that may provide the basis of diverse behaviors, including sensory perception, motor integration, working memory, and attention.
Figures
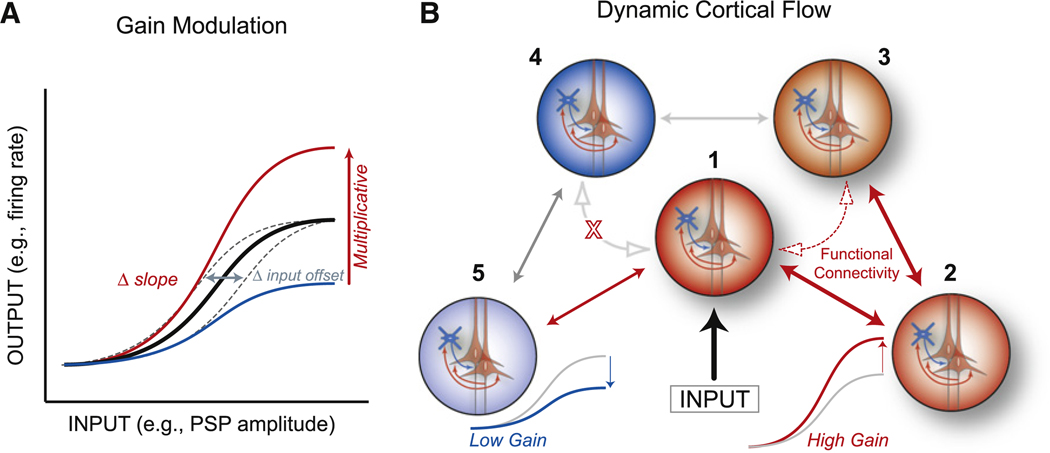
(A) Schematic of various forms of response modulation. Input gain (dashed gray) shifts the threshold and saturation points without altering the ratio of input to output. Multiplicative gain increases (red) or divisively decreases (blue) the responsiveness at all levels of input by a similar percentage and results in a change in the slope of the input-output function. (B) Schematic diagram illustrating five sets of neurons or neuronal groups that are anatomically interconnected (double-headed arrows), with the middle group (1) receiving an input. If neuronal group2 has high gain, then groups 1, 2, and 3 exhibit enhanced functional connectivity (dashed arrows), with activity flowing easily between groups 1 and 3. Likewise, if group 5 has low gain, then the flow of information and interactions between groups 1, 4, and 5 will be limited. Note that the responsiveness (gain) within and among neuronal groups is determined by local recurrent excitation and inhibition.

Illustrated are simultaneous recordings of the extracellular local field potential (LFP, top), the multiple unit activity (MU, middle), and intracellular membrane potential (bottom) from a cortical pyramidal cell during the generation of one Up state (bordered by two Down states). Note that the Up state is associated with a marked increase in local network activity, depolarization of the neuron (~25 mV), a marked increase in membrane potential variance (SD of Up = 2.5 mV), and the presence of higher-frequency (gamma, ~40 Hz) oscillations in the LFP. Periods of depolarization mediated by network activity provide “windows of opportunity” based upon increased neuronal gain and may be of long (example 1), medium (example 2), or short (example 3) duration. Thus, various frequency components of synaptic activity interact to initiate action potentials. Inset expands window 3, where the depolarizing half (~12.5 ms) of one full LFP oscillation cycle provides a short temporal window for the integration of both excitatory and inhibitory postsynaptic potentials (PSPs).

(A) In the absence of neuronal variance, neurons respond to depolarization with an abrupt increase in firing rate once threshold has been reached (black). However, in the presence of membrane potential variance, the average relationship between firing rate and membrane potential can exhibit a power law (red). (B) Intracellular recordings of cortical neuronal responses to visual stimuli in vivo have revealed, on average, a power law relationship between membrane potential and firing rate. (C) In the presence of a power law relationship between membrane potential and firing rate, changes in either excitatory or inhibitory background conductance that depolarize or hyperpolarize neurons (±2–5 mV), respectively, can result in multiplicative-like changes in the input-output relationship of cortical neurons. (D) Depolarization induced through the intracellular injection of current results in a multiplicative-like change in the contrast response function curve of visual cortical neurons. (E) Scaling the hyperpolarized curve from (D) with a constant gain factor reveals a multiplicative-like gain change. (F) Depolarization that occurs spontaneously owing to synaptic bombardment in vivo can also result in multiplicative-like increases in neuronal responsiveness of the contrast response function. (B) Adapted by permission from Macmillan Publishers Ltd: Nature Neuroscience (Priebe et al., 2004), copyright 2004; (C) modified from Murphy and Miller (2003); (D) modified from Sanchez-Vives et al. (2000); (E) modified from Haider et al. (2007).

(A) Short-latency (putative monosynaptic) neuronal interactions between simultaneously recorded putative pyramidal (red) and inhibitory (blue) cells in rat prefrontal cortex during the performance of a maze task, as judged from extracellular recordings and cross-correlation spike histograms. (B) For this pair, the strength of interaction varied as a function of spatial position in the maze, with the strongest interactions apparent near the decision point of the maze (middle). (C) Local network diagram constructed from ensembles of simultaneously recorded pyramidal and putative inhibitory neurons reveal similar short-latency enhancement of monosynaptic interactions that are spatially dependent and whose strength varies significantly with behavioral demands. Interactions in red (1, 2) are significantly enhanced on rightward turns during maze running. (D) Same ensemble of neurons upon leftward turns during maze running. Some functional connections that are spatially modulated are present in both directions of running (2), while others appear (3) or disappear (1) during leftward turns, indicating rapid changes in functional connectivity during ongoing behavior. Adapted by permission from Macmillan Publishers Ltd: Nature Neuroscience (Fujisawa et al., 2008), copyright 2008.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
