

Interplay between cytosolic dopamine, calcium, and alpha-synuclein causes selective death of substantia nigra neurons
- PMID: 19409267
- PMCID: PMC2677560
- DOI: 10.1016/j.neuron.2009.01.033
Interplay between cytosolic dopamine, calcium, and alpha-synuclein causes selective death of substantia nigra neurons
Abstract
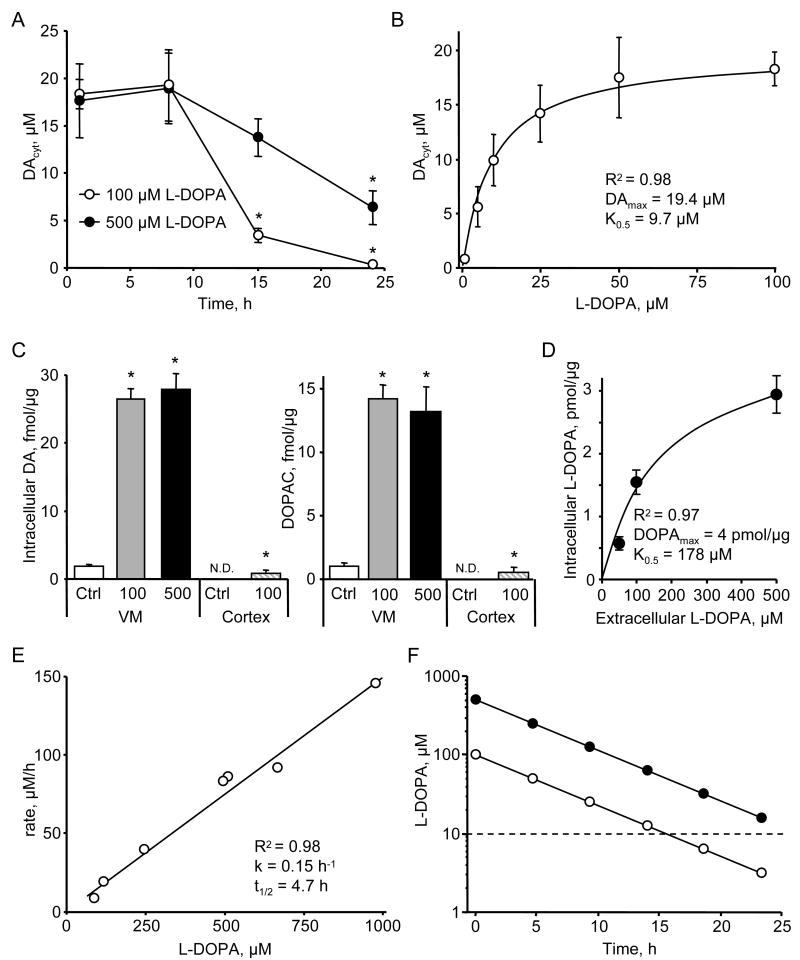
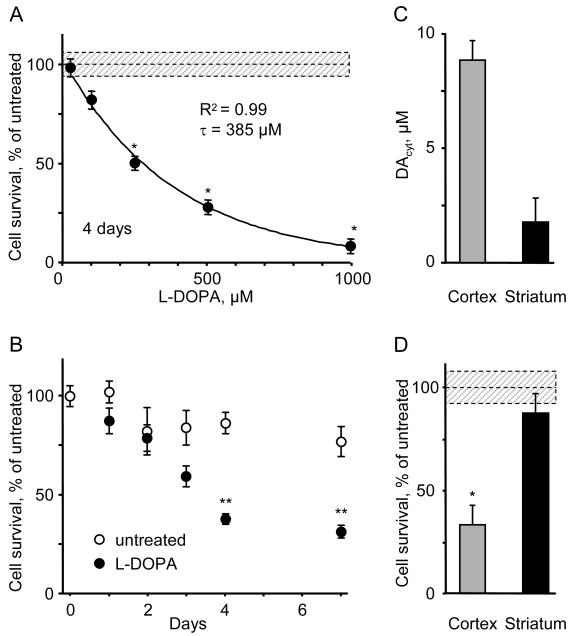
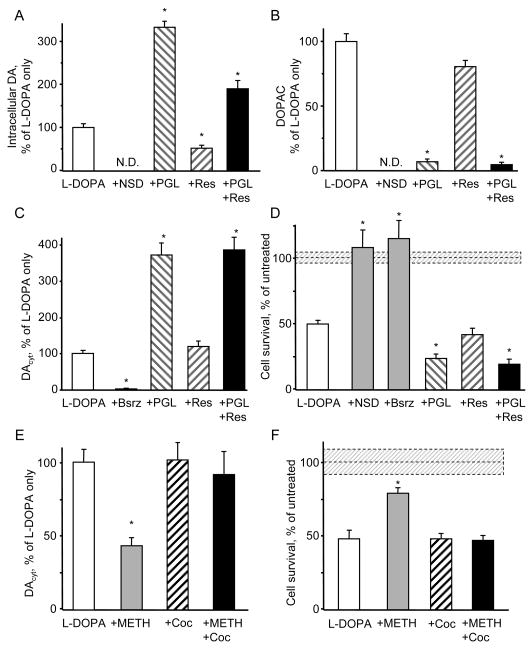
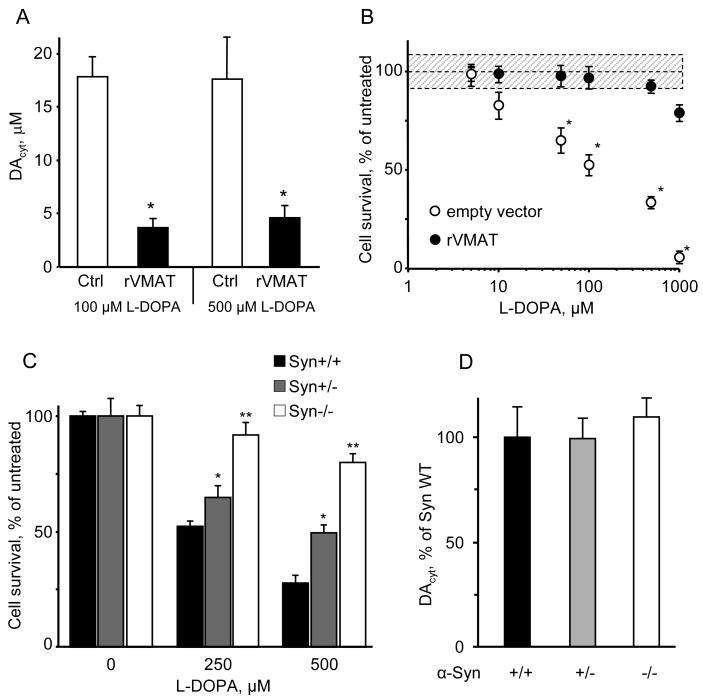
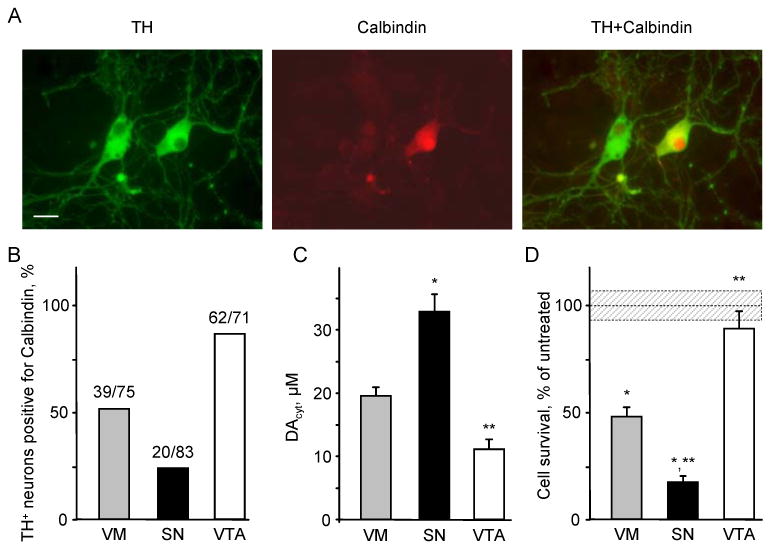
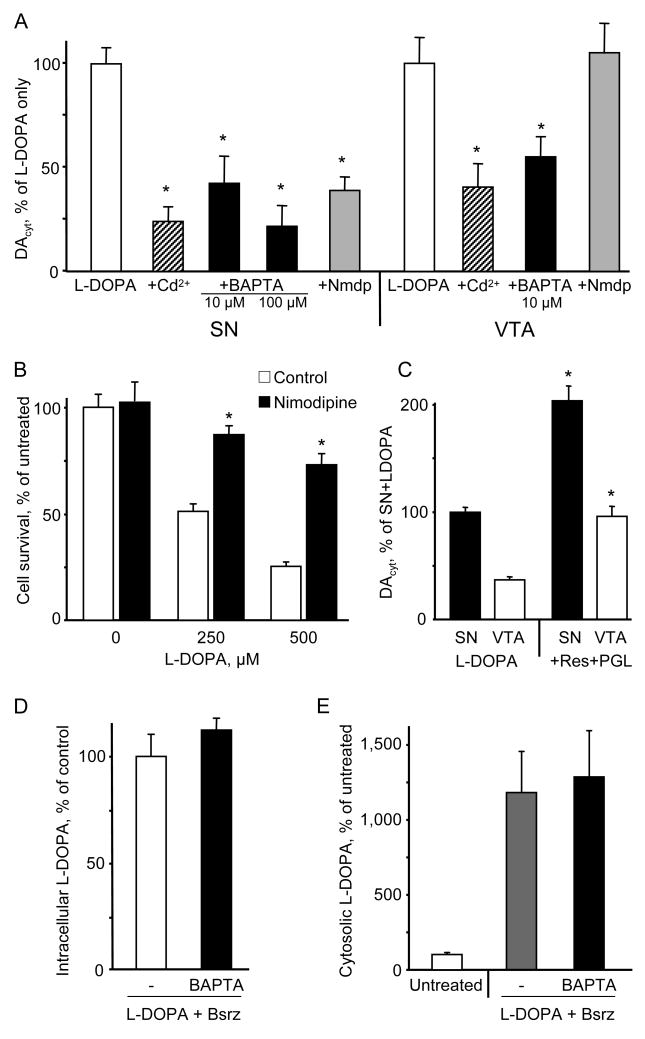
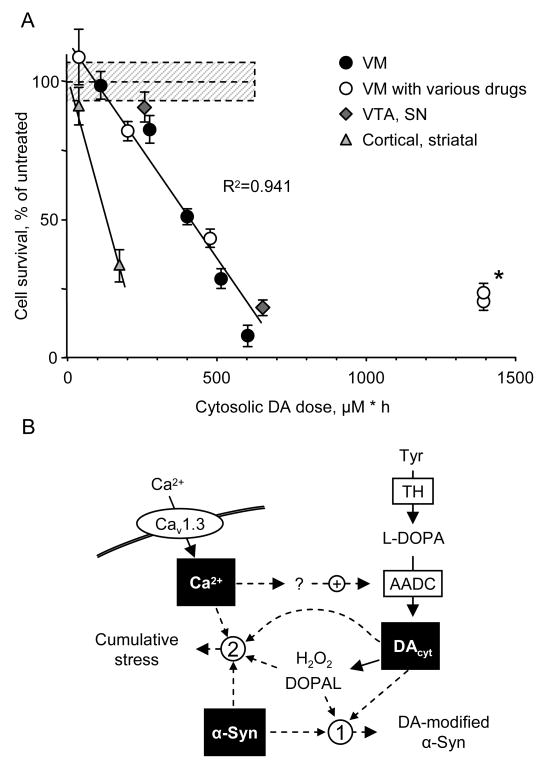
The basis for selective death of specific neuronal populations in neurodegenerative diseases remains unclear. Parkinson's disease (PD) is a synucleinopathy characterized by a preferential loss of dopaminergic neurons in the substantia nigra (SN), whereas neurons of the ventral tegmental area (VTA) are spared. Using intracellular patch electrochemistry to directly measure cytosolic dopamine (DA(cyt)) in cultured midbrain neurons, we confirm that elevated DA(cyt) and its metabolites are neurotoxic and that genetic and pharmacological interventions that decrease DA(cyt) provide neuroprotection. L-DOPA increased DA(cyt) in SN neurons to levels 2- to 3-fold higher than in VTA neurons, a response dependent on dihydropyridine-sensitive Ca2+ channels, resulting in greater susceptibility of SN neurons to L-DOPA-induced neurotoxicity. DA(cyt) was not altered by alpha-synuclein deletion, although dopaminergic neurons lacking alpha-synuclein were resistant to L-DOPA-induced cell death. Thus, an interaction between Ca2+, DA(cyt), and alpha-synuclein may underlie the susceptibility of SN neurons in PD, suggesting multiple therapeutic targets.
Figures
Comment in
-
A lethal convergence of dopamine and calcium.Neuron. 2009 Apr 30;62(2):163-4. doi: 10.1016/j.neuron.2009.04.010. Neuron. 2009. PMID: 19409260
References
-
- Abeliovich A, Schmitz Y, Farinas I, Choi-Lundberg D, Ho WH, Castillo PE, Shinsky N, Verdugo JM, Armanini M, Ryan A, et al. Mice lacking alpha-synuclein display functional deficits in the nigrostriatal dopamine system. Neuron. 2000;25:239–252. - PubMed
-
- Abou-Sleiman PM, Muqit MM, Wood NW. Expanding insights of mitochondrial dysfunction in Parkinson's disease. Nat Rev Neurosci. 2006;7:207–219. - PubMed
-
- Alvarez-Fischer D, Henze C, Strenzke C, Westrich J, Ferger B, Hoglinger GU, Oertel WH, Hartmann A. Characterization of the striatal 6-OHDA model of Parkinson's disease in wild type and alpha-synuclein-deleted mice. Exp Neurol. 2008;210:182–193. Epub 2007 Nov 2001. - PubMed
-
- Baroudi G, Qu Y, Ramadan O, Chahine M, Boutjdir M. Protein kinase C activation inhibits Cav1.3 calcium channel at NH2-terminal serine 81 phosphorylation site. Am J Physiol Heart Circ Physiol. 2006;291:H1614–1622. - PubMed
-
- Burke RE, Antonelli M, Sulzer D. Glial cell line-derived neurotrophic growth factor inhibits apoptotic death of postnatal substantia nigra dopamine neurons in primary culture. J Neurochem. 1998;71:517–525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
