

Centromere-specific assembly of CENP-a nucleosomes is mediated by HJURP
- PMID: 19410544
- PMCID: PMC2747366
- DOI: 10.1016/j.cell.2009.02.039
Centromere-specific assembly of CENP-a nucleosomes is mediated by HJURP
Abstract
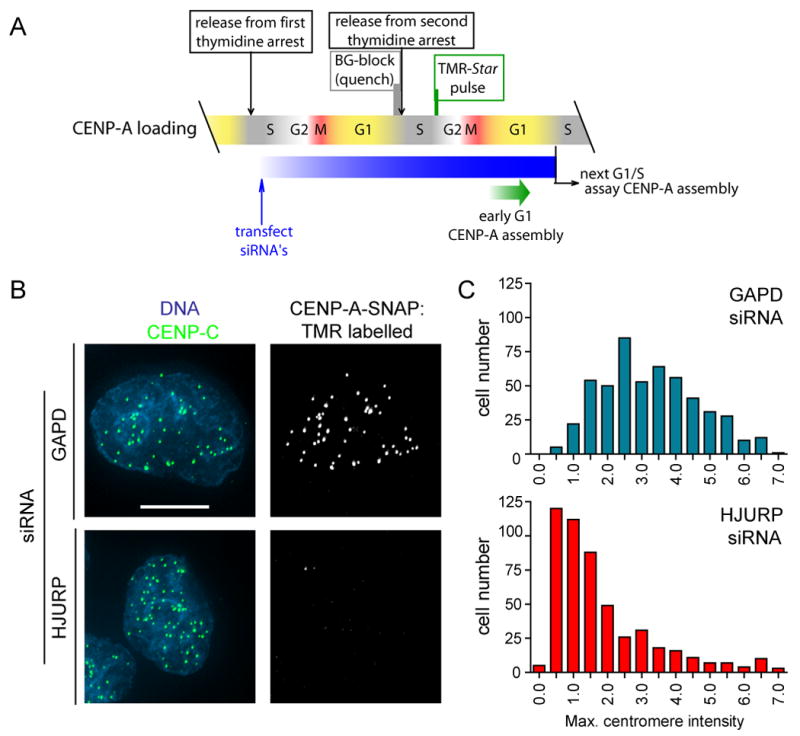
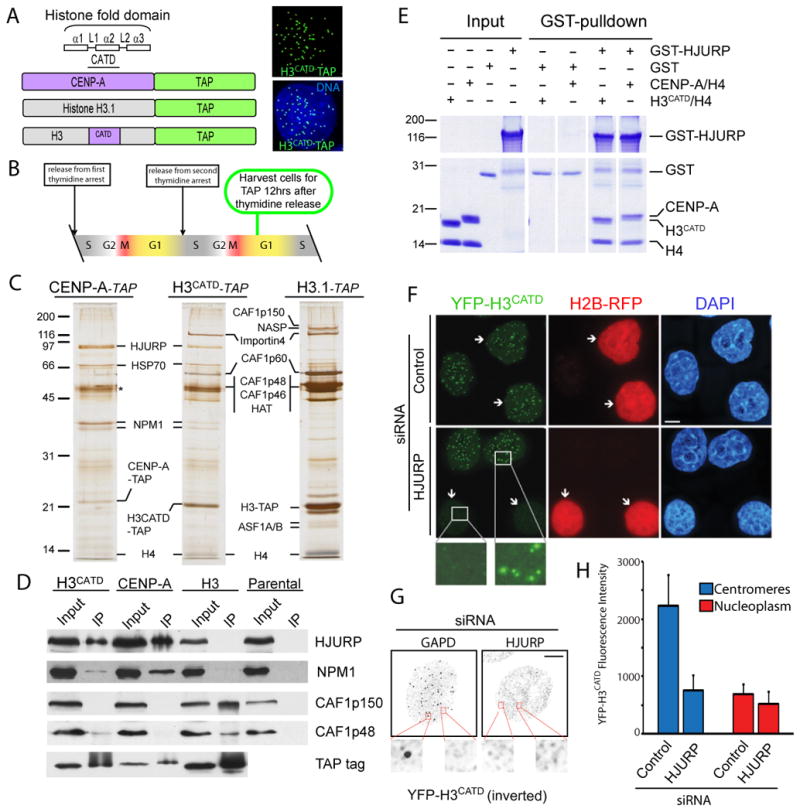
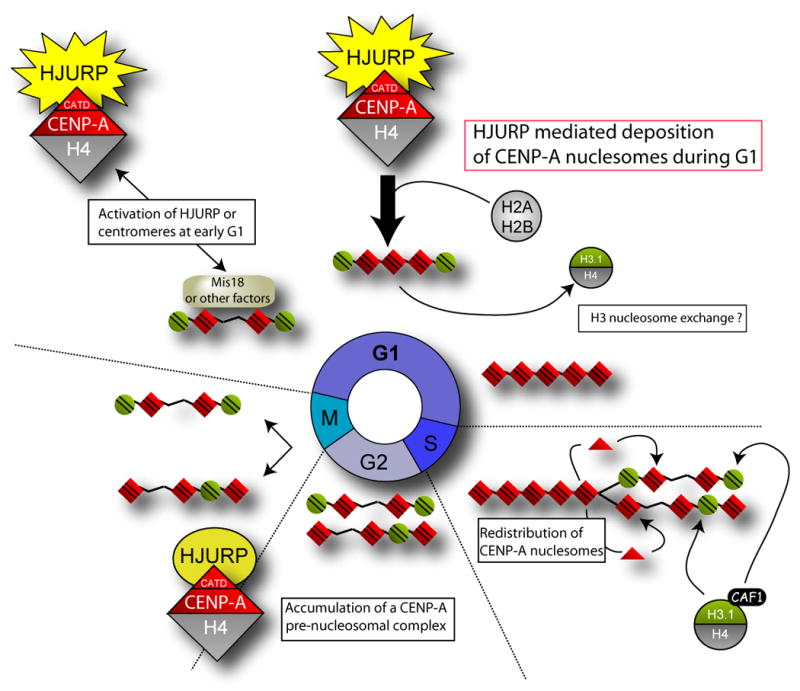
The centromere is responsible for accurate chromosome segregation. Mammalian centromeres are specified epigenetically, with all active centromeres containing centromere-specific chromatin in which CENP-A replaces histone H3 within the nucleosome. The proteins responsible for assembly of human CENP-A into centromeric nucleosomes during the G1 phase of the cell cycle are shown here to be distinct from the chromatin assembly factors previously shown to load other histone H3 variants. Here we demonstrate that prenucleosomal CENP-A is complexed with histone H4, nucleophosmin 1, and HJURP. Recruitment of new CENP-A into nucleosomes at replicated centromeres is dependent on HJURP. Recognition by HJURP is mediated through the centromere targeting domain (CATD) of CENP-A, a region that we demonstrated previously to induce a unique conformational rigidity to both the subnucleosomal CENP-A heterotetramer and the corresponding assembled nucleosome. We propose HJURP to be a cell-cycle-regulated CENP-A-specific histone chaperone required for centromeric chromatin assembly.
Figures
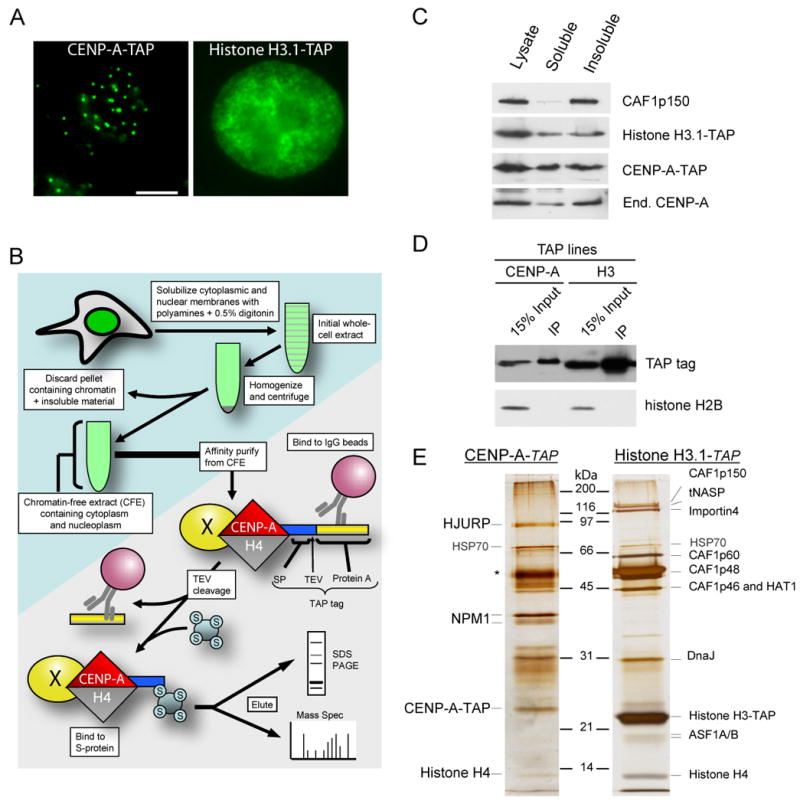
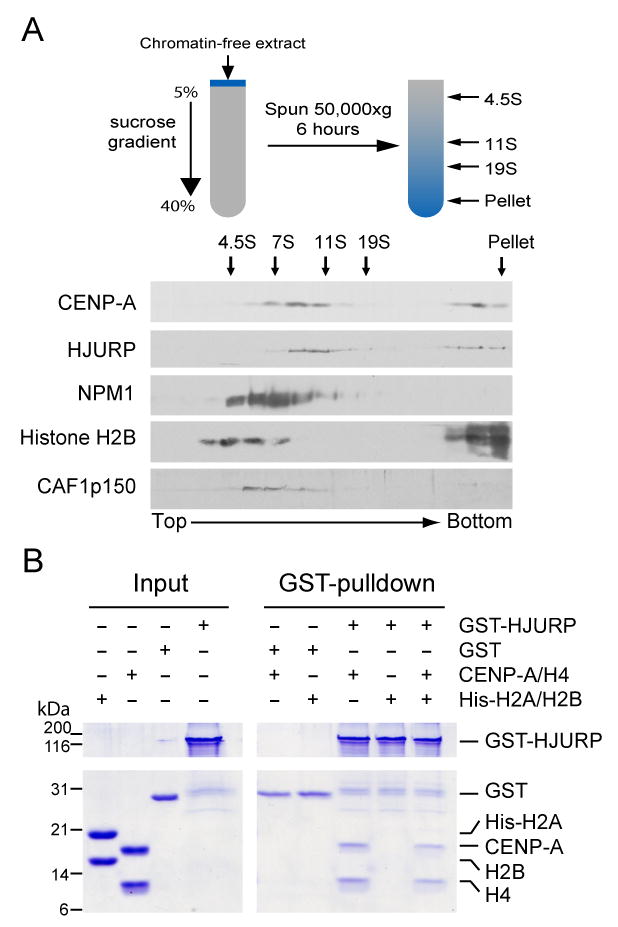
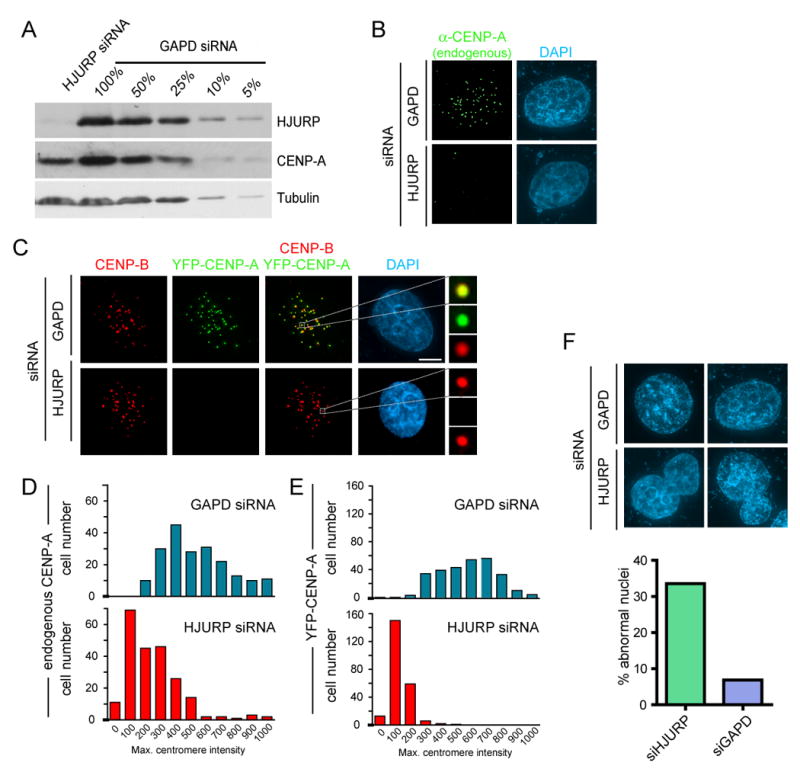
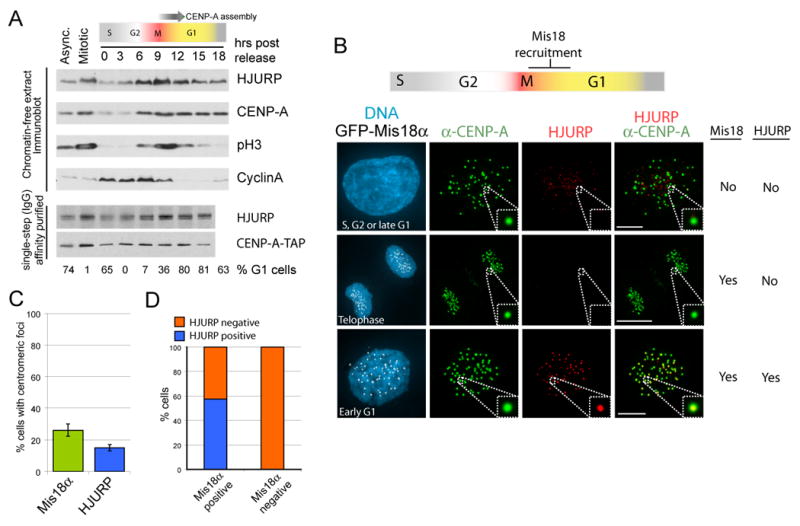
References
-
- Ahmad K, Henikoff S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol Cell. 2002;9:1191–1200. - PubMed
-
- Black BE, Foltz DR, Chakravarthy S, Luger K, Woods VL, Jr, Cleveland DW. Structural determinants for generating centromeric chromatin. Nature. 2004;430:578–582. - PubMed
-
- Black BE, Jansen LE, Maddox PS, Foltz DR, Desai AB, Shah JV, Cleveland DW. Centromere identity maintained by nucleosomes assembled with histone H3 containing the CENP-A targeting domain. Mol Cell. 2007;25:309–322. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
