

Role of protein phosphatases and mitochondria in the neuroprotective effects of estrogens
- PMID: 19410596
- PMCID: PMC2835549
- DOI: 10.1016/j.yfrne.2009.04.013
Role of protein phosphatases and mitochondria in the neuroprotective effects of estrogens
Abstract
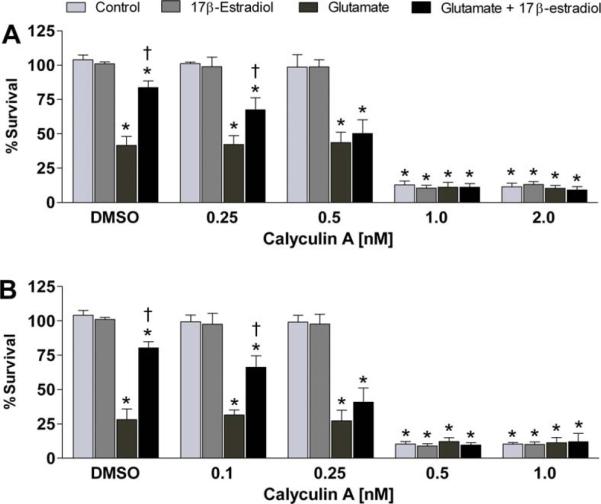
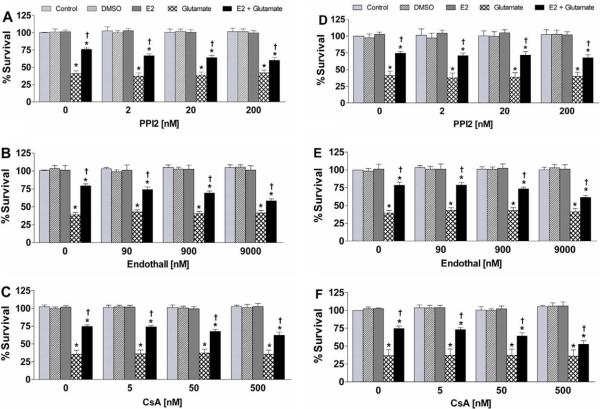
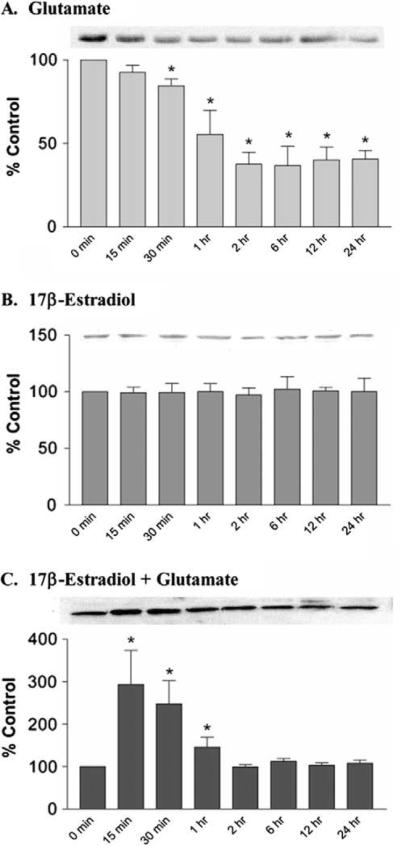
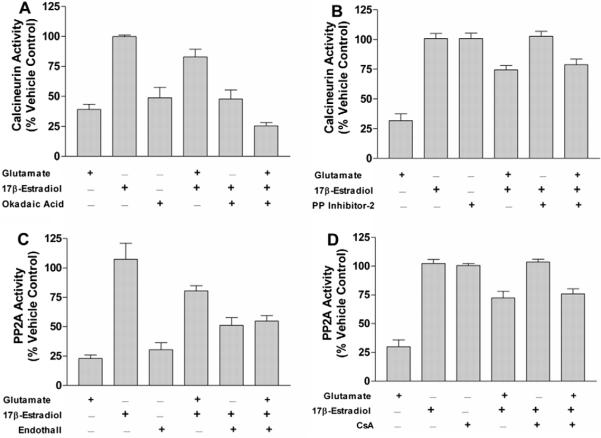
In the present treatise, we provide evidence that the neuroprotective and mito-protective effects of estrogens are inexorably linked and involve the ability of estrogens to maintain mitochondrial function during neurotoxic stress. This is achieved by the induction of nuclear and mitochondrial gene expression, the maintenance of protein phosphatases levels in a manner that likely involves modulation of the phosphorylation state of signaling kinases and mitochondrial pro- and anti-apoptotic proteins, and the potent redox/antioxidant activity of estrogens. These estrogen actions are mediated through a combination of estrogens receptor (ER)-mediated effects on nuclear and mitochondrial transcription of protein vital to mitochondrial function, ER-mediated, non-genomic signaling and non-ER-mediated effects of estrogens on signaling and oxidative stress. Collectively, these multifaceted, coordinated action of estrogens leads to their potency in protecting neurons from a wide variety of acute insults as well as chronic neurodegenerative processes.
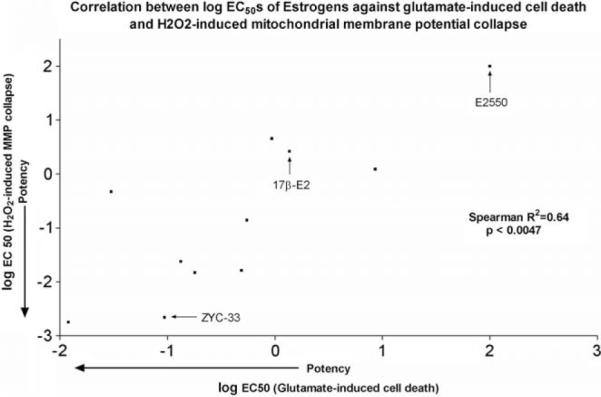
Figures
Similar articles
-
Role of protein phosphatases in estrogen-mediated neuroprotection.J Neurosci. 2005 Aug 3;25(31):7191-8. doi: 10.1523/JNEUROSCI.1328-05.2005. J Neurosci. 2005. PMID: 16079401 Free PMC article.
-
Estrogen receptor-independent neuroprotection via protein phosphatase preservation and attenuation of persistent extracellular signal-regulated kinase 1/2 activation.J Pharmacol Exp Ther. 2008 Mar;324(3):1188-95. doi: 10.1124/jpet.107.132308. Epub 2007 Dec 18. J Pharmacol Exp Ther. 2008. PMID: 18089844
-
Mitochondria as therapeutic targets of estrogen action in the central nervous system.Curr Drug Targets CNS Neurol Disord. 2004 Aug;3(4):297-313. doi: 10.2174/1568007043337193. Curr Drug Targets CNS Neurol Disord. 2004. PMID: 15379606 Review.
-
Neuroprotection by estrogen in the brain: the mitochondrial compartment as presumed therapeutic target.J Neurochem. 2009 Jul;110(1):1-11. doi: 10.1111/j.1471-4159.2009.06133.x. Epub 2009 Apr 29. J Neurochem. 2009. PMID: 19457121 Review.
-
Protein phosphatase 1, protein phosphatase 2A, and calcineurin play a role in estrogen-mediated neuroprotection.Endocrinology. 2008 Oct;149(10):5235-43. doi: 10.1210/en.2008-0610. Epub 2008 Jun 19. Endocrinology. 2008. PMID: 18566123 Free PMC article.
Cited by
-
Postmortem mitochondrial membrane permeability transition assessment of apoptotic cell death in brain and liver of insulin resistant, ovariectomised rats.IBRO Neurosci Rep. 2021 Oct 9;11:156-163. doi: 10.1016/j.ibneur.2021.09.004. eCollection 2021 Dec. IBRO Neurosci Rep. 2021. PMID: 34939064 Free PMC article.
-
Estrogen protection in Friedreich's ataxia skin fibroblasts.Endocrinology. 2011 Jul;152(7):2742-9. doi: 10.1210/en.2011-0184. Epub 2011 May 3. Endocrinology. 2011. PMID: 21540287 Free PMC article.
-
Early and gender-specific differences in spinal cord mitochondrial function and oxidative stress markers in a mouse model of ALS.Acta Neuropathol Commun. 2016 Jan 13;4:3. doi: 10.1186/s40478-015-0271-6. Acta Neuropathol Commun. 2016. PMID: 26757991 Free PMC article.
-
Protective Action of Neurotrophic Factors and Estrogen against Oxidative Stress-Mediated Neurodegeneration.J Toxicol. 2011;2011:405194. doi: 10.1155/2011/405194. Epub 2011 May 31. J Toxicol. 2011. PMID: 21776259 Free PMC article.
-
Genetic dissection of strain dependent paraquat-induced neurodegeneration in the substantia nigra pars compacta.PLoS One. 2012;7(1):e29447. doi: 10.1371/journal.pone.0029447. Epub 2012 Jan 24. PLoS One. 2012. PMID: 22291891 Free PMC article.
References
-
- Akita Y, Kawasaki H, Imajoh-Ohmi S, Fukuda H, Ohno S, Hirano H, Ono Y, Yonekawa H. Protein kinase C epsilon phosphorylates keratin 8 at Ser8 and Ser23 in GH4C1 cells stimulated by thyrotropin-releasing hormone. FEBS J. 2007;274:3270–3285. - PubMed
-
- Alkayed NJ, Harukuni I, Kimes AS, London ED, Traystman RJ, Hurn PD. Gender-linked brain injury in experimental stroke. Stroke. 1998;29:159–165. discussion 166. - PubMed
-
- Arendt T, Holzer M, Fruth R, Bruckner MK, Gartner U. Paired helical filament-like phosphorylation of tau, deposition of beta/A4-amyloid and memory impairment in rat induced by chronic inhibition of phosphatase 1 and 2A. Neuroscience. 1995;69:691–698. - PubMed
-
- Arias C, Sharma N, Davies P, Shafit-Zagardo B. Okadaic acid induces early changes in microtubule-associated protein 2 and tau phosphorylation prior to neurodegeneration in cultured cortical neurons. J. Neurochem. 1993;61:673–682. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
