

Meiotic recombination generates rich diversity in NK cell receptor genes, alleles, and haplotypes
- PMID: 19411600
- PMCID: PMC2675964
- DOI: 10.1101/gr.085738.108
Meiotic recombination generates rich diversity in NK cell receptor genes, alleles, and haplotypes
Abstract
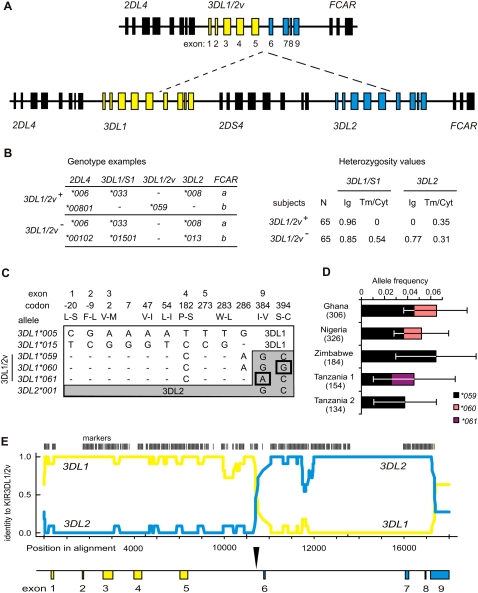
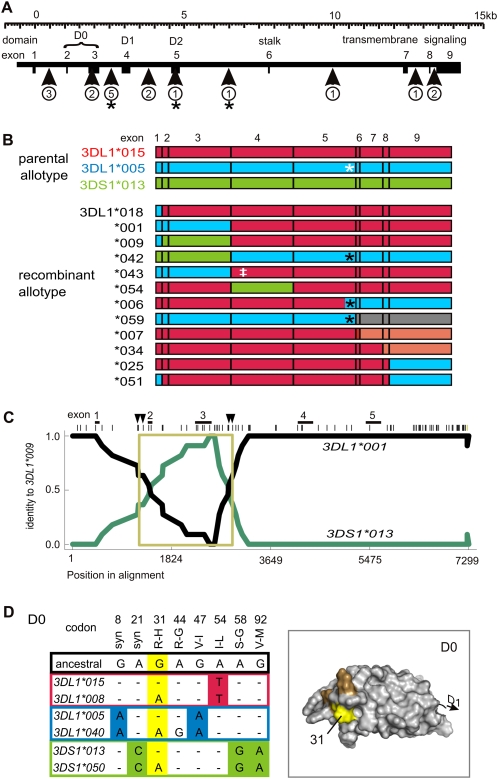
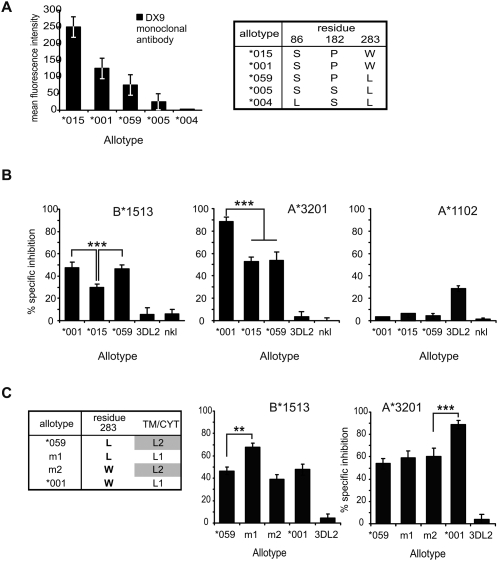
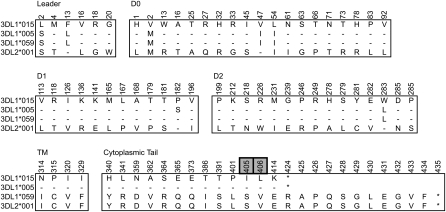
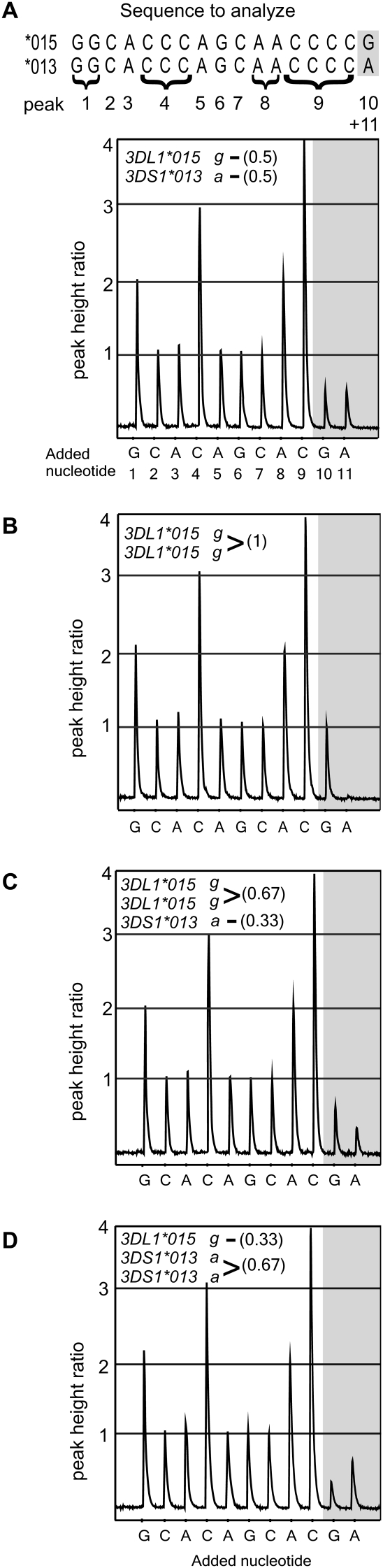
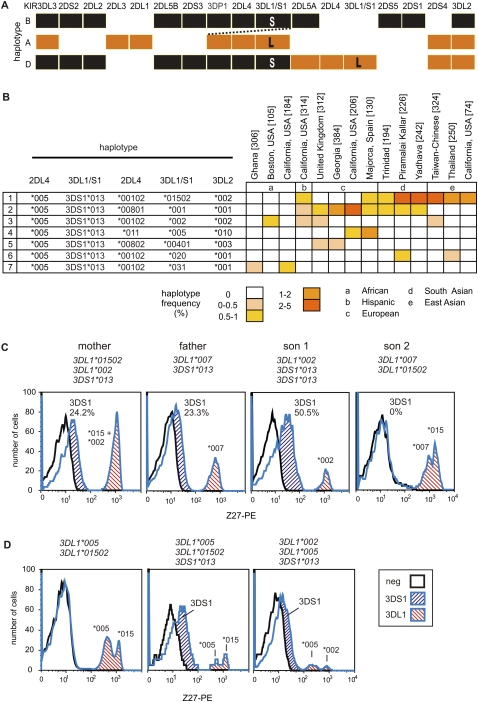
Natural killer (NK) cells contribute to the essential functions of innate immunity and reproduction. Various genes encode NK cell receptors that recognize the major histocompatibility complex (MHC) Class I molecules expressed by other cells. For primate NK cells, the killer-cell immunoglobulin-like receptors (KIR) are a variable and rapidly evolving family of MHC Class I receptors. Studied here is KIR3DL1/S1, which encodes receptors for highly polymorphic human HLA-A and -B and comprises three ancient allelic lineages that have been preserved by balancing selection throughout human evolution. While the 3DS1 lineage of activating receptors has been conserved, the two 3DL1 lineages of inhibitory receptors were diversified through inter-lineage recombination with each other and with 3DS1. Prominent targets for recombination were D0-domain polymorphisms, which modulate enhancer function, and dimorphism at position 283 in the D2 domain, which influences inhibitory function. In African populations, unequal crossing over between the 3DL1 and 3DL2 genes produced a deleted KIR haplotype in which the telomeric "half" was reduced to a single fusion gene with functional properties distinct from its 3DL1 and 3DL2 parents. Conversely, in Eurasian populations, duplication of the KIR3DL1/S1 locus by unequal crossing over has enabled individuals to carry and express alleles of all three KIR3DL1/S1 lineages. These results demonstrate how meiotic recombination combines with an ancient, preserved diversity to create new KIR phenotypes upon which natural selection acts. A consequence of such recombination is to blur the distinction between alleles and loci in the rapidly evolving human KIR gene family.
Figures
References
-
- Alvarez-Arias D.A., Campbell K.S. Protein kinase C regulates expression and function of inhibitory killer cell Ig-like receptors in NK cells. J. Immunol. 2007;179:5281–5290. - PubMed
-
- Artavanis-Tsakonas K., Eleme K., McQueen K.L., Cheng N.W., Parham P., Davis D.M., Riley E.M. Activation of a subset of human NK cells upon contact with Plasmodium falciparum-infected erythrocytes. J. Immunol. 2003;171:5396–5405. - PubMed
-
- Bailey J.A., Eichler E.E. Primate segmental duplications: Crucibles of evolution, diversity and disease. Nat. Rev. Genet. 2006;7:552–564. - PubMed
-
- Bashirova A.A., Martin M.P., McVicar D.W., Carrington M. The killer immunoglobulin-like receptor gene cluster: Tuning the genome for defense. Annu. Rev. Genomics Hum. Genet. 2006;7:277–300. - PubMed
-
- Belle I., Hou L., Chen M., Steiner N.K., Ng J., Hurley C.K. Investigation of killer cell immunoglobulin-like receptor gene diversity in KIR3DL1 and KIR3DS1 in a transplant population. Tissue Antigens. 2008;71:434–439. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials