

Hexameric assembly of the proteasomal ATPases is templated through their C termini
- PMID: 19412160
- PMCID: PMC2722381
- DOI: 10.1038/nature08065
Hexameric assembly of the proteasomal ATPases is templated through their C termini
Abstract
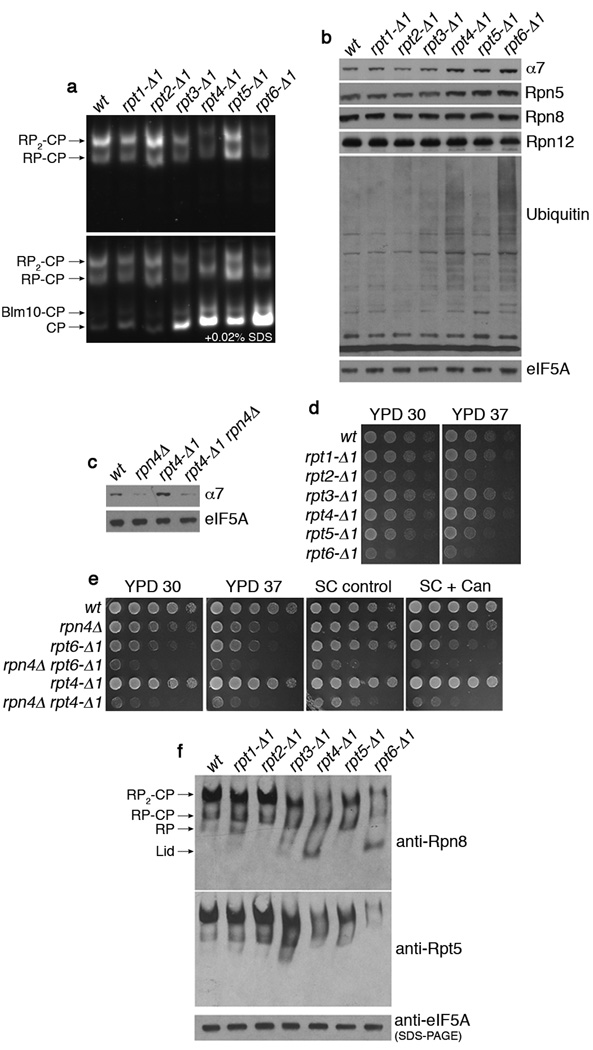
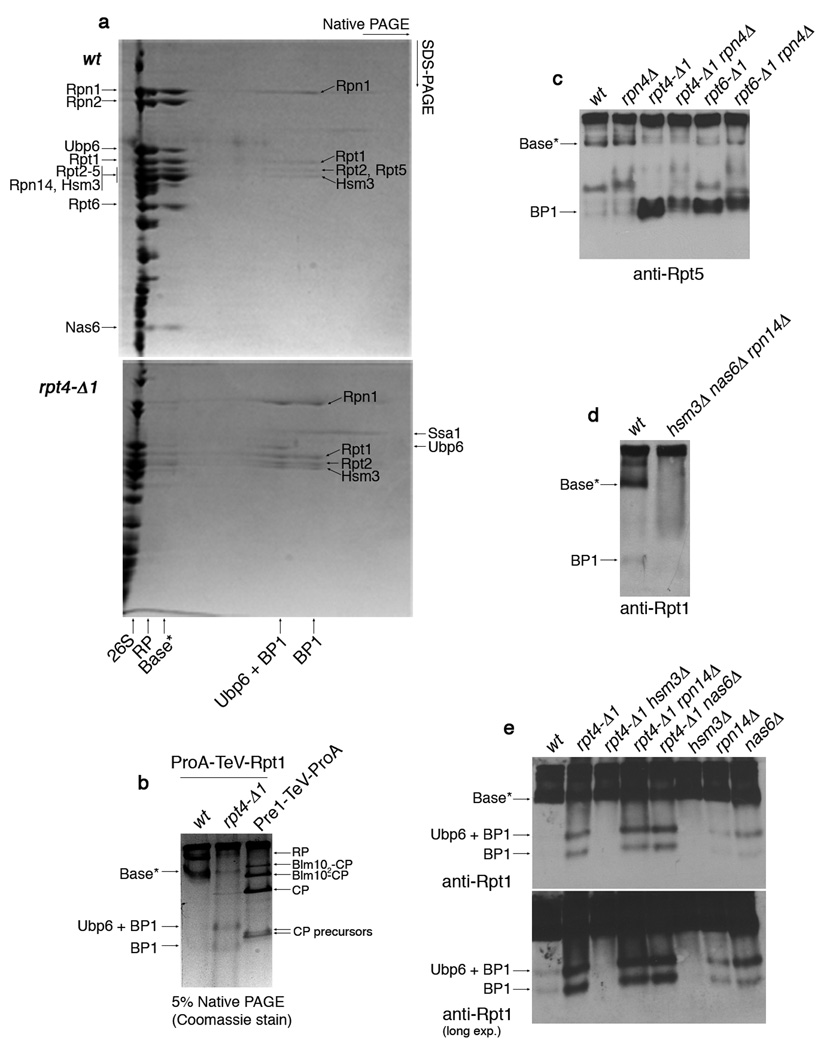
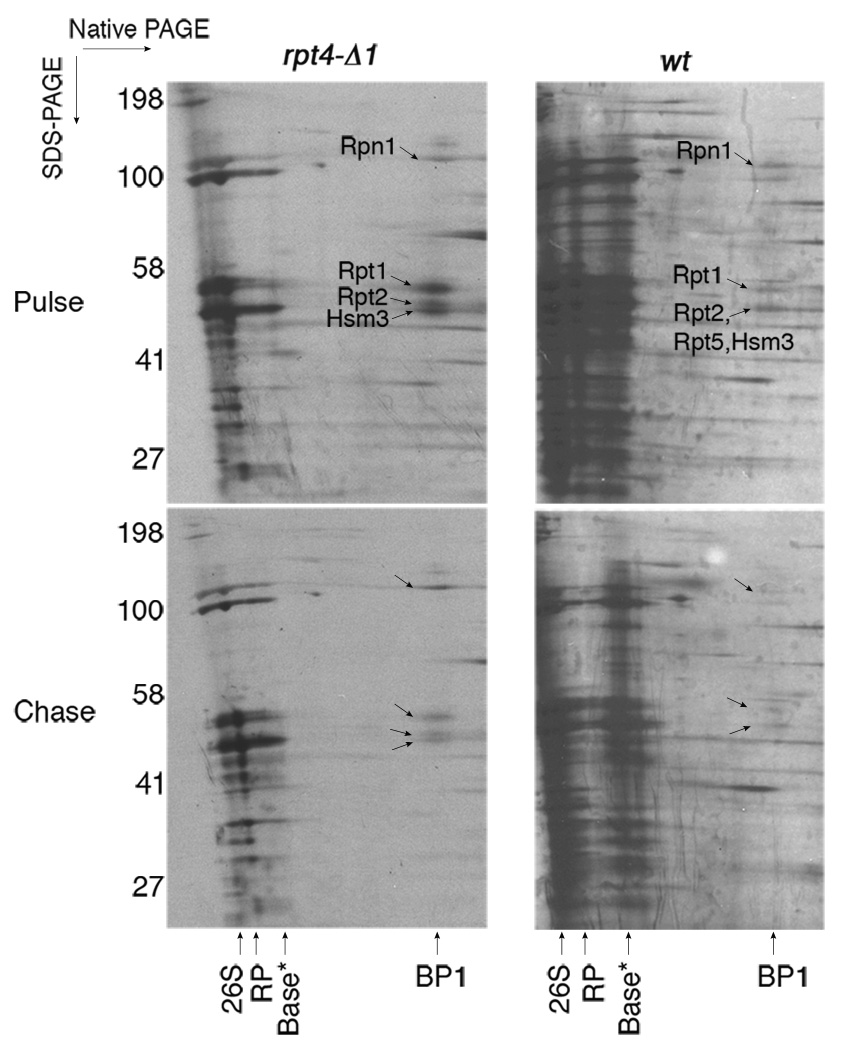
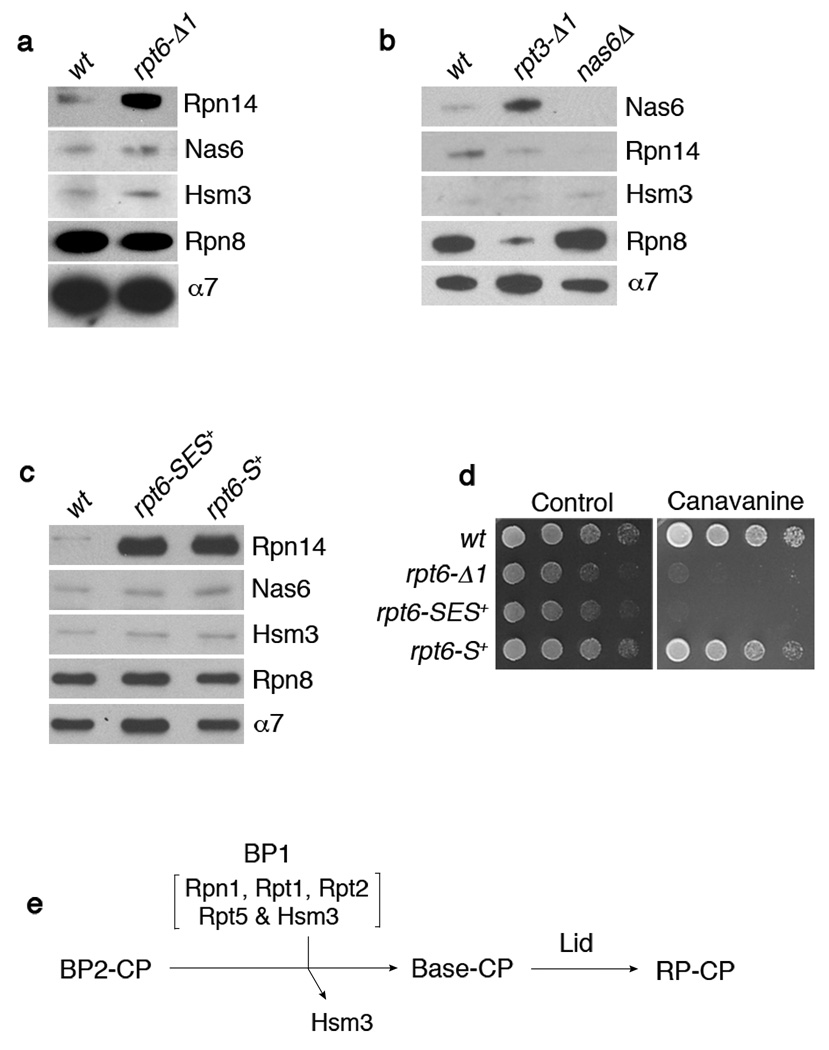
Substrates of the proteasome are recognized and unfolded by the regulatory particle, and then translocated into the core particle (CP) to be degraded. A hetero-hexameric ATPase ring, containing subunits Rpt1-6, is situated within the base subassembly of the regulatory particle. The ATPase ring sits atop the CP, with the Rpt carboxy termini inserted into pockets in the CP. Here we identify a previously unknown function of the Rpt proteins in proteasome biogenesis through deleting the C-terminal residue from each Rpt in the yeast Saccharomyces cerevisiae. Our results indicate that assembly of the hexameric ATPase ring is templated on the CP. We have also identified an apparent intermediate in base assembly, BP1, which contains Rpn1, three Rpts and Hsm3, a chaperone for base assembly. The Rpt proteins with the strongest assembly phenotypes, Rpt4 and Rpt6, were absent from BP1. We propose that Rpt4 and Rpt6 form a nucleating complex to initiate base assembly, and that this complex is subsequently joined by BP1 to complete the Rpt ring. Our studies show that assembly of the proteasome base is a rapid yet highly orchestrated process.
Figures
Comment in
-
Cell biology: The proteasome assembly line.Nature. 2009 Jun 11;459(7248):787-8. doi: 10.1038/459787a. Nature. 2009. PMID: 19516331 Free PMC article.
References
-
- Forster A, Masters EI, Whitby FG, Robinson H, Hill CP. The 1.9 A structure of a proteasome-11S activator complex and implications for proteasome-PAN/PA700 interactions. Mol. Cell. 2005;18:589–599. - PubMed
-
- Whitby FG, et al. Structural basis for the activation of 20S proteasomes by 11S regulators. Nature. 2000;408:115–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
