

Enzymatic characterization of a human acyltransferase activity
- PMID: 19412546
- PMCID: PMC2672172
- DOI: 10.1371/journal.pone.0005426
Enzymatic characterization of a human acyltransferase activity
Abstract
Background: Non-histone protein acylation is increasingly recognized as an important posttranslational modification, but little is known as to the biochemical properties of protein serine acylating enzymes.
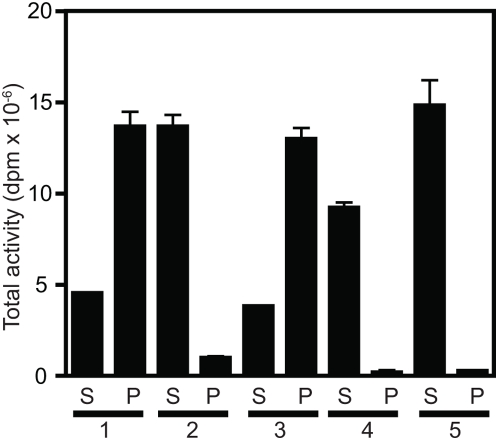
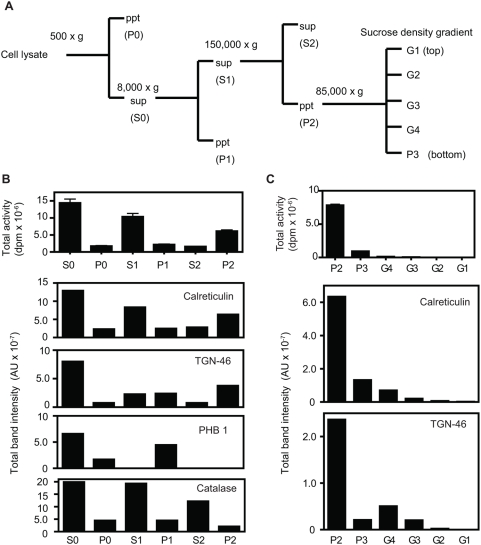
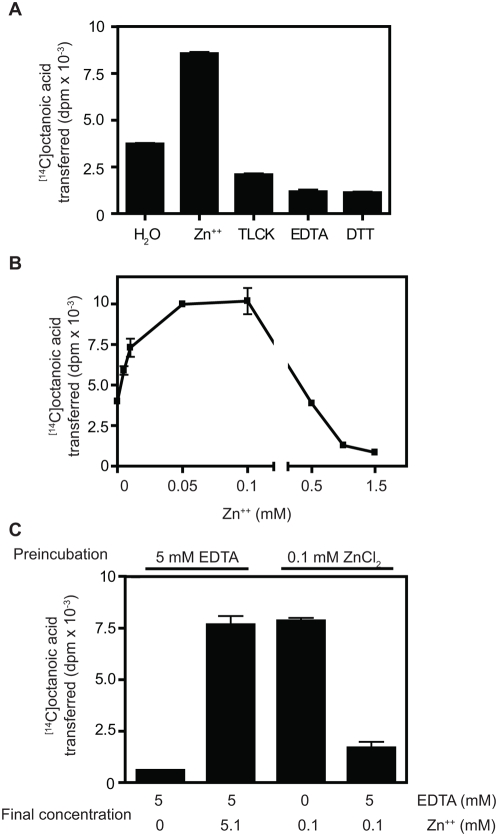
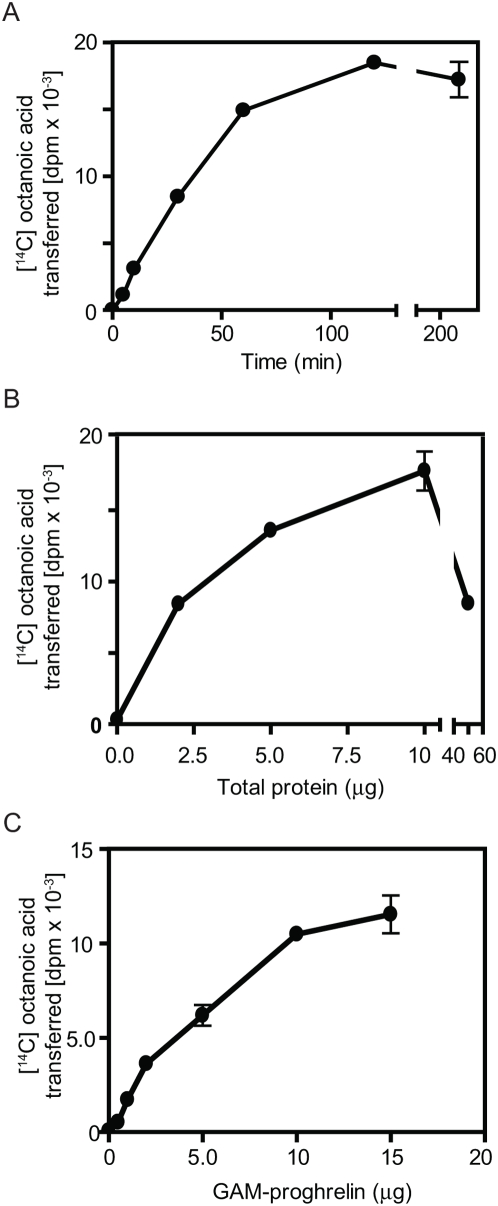
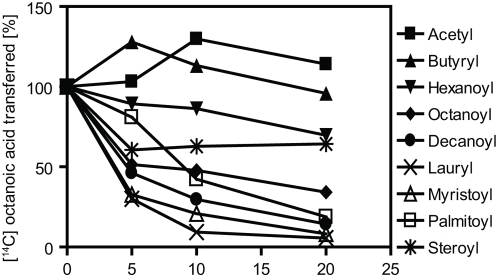
Methodology/principal findings: We here report that we have identified a metal-stimulated serine octanoyltransferase activity in microsomes from human erythroleukemic (HEL) cells. The HEL acylating enzyme was linear with respect to time and protein, exhibited a neutral pH optimum (stimulated by cobalt and zinc), and inhibited by chelating reagents. Hydroxylamine treatment removed most, but not all, of the attached radioactivity. A salt extract of microsomal membranes contained the major portion of enzyme activity, indicating that this acyltransferase is not an integral membrane protein. Sucrose density fractionation showed that the acyltransferase activity is concentrated in the endoplasmic reticulum. In competition experiments, the acyltransferase was well inhibited by activated forms of fatty acids containing at least eight to fourteen carbons, but not by acetyl CoA. The zinc-stimulated HEL acyltransferase did not octanoylate proenkephalin, proopiomelanocortin, His-tagged proghrelin, or proghrelin lacking the amino-terminal His-tag stub of Gly-Ala-Met. The peptides des-acyl ghrelin and ACTH were also not acylated; however, des-acyl ghrelin containing the N-terminal tripeptide Gly-Ala-Met was acylated. Mutagenesis studies indicated a requirement for serine five residues from the amino terminus, reminiscent of myristoyl transferase, but not of ghrelin acylation. However, recombinant myristoyl transferase could not recapitulate the hydroxylamine sensitivity, zinc-stimulation, nor EDTA inhibition obtained with HEL acyltransferase, properties preserved in the HEL cell enzyme purified through four sequential chromatographic steps.
Conclusions/significance: In conclusion, our data demonstrate the presence of a zinc-stimulated acyltransferase activity concentrated in the endoplasmic reticulum in HEL cells which is likely to contribute to medium-chain protein lipidation.
Conflict of interest statement
Figures



References
-
- Wilkinson CW. Roles of acetylation and other post-translational modifications in melanocortin function and interactions with endorphins. Peptides. 2006;27:453–71. - PubMed
-
- van der Lely AJ, Tschop M, Heiman ML, Ghigo E. Biological, physiological, pathophysiological, and pharmacological aspects of ghrelin. Endocr Rev. 2004;25:426–57. - PubMed
-
- Kojima M, Kangawa K. Ghrelin: structure and function. Physiol Rev. 2005;85:495–522. - PubMed
-
- Mukherjee S, Keitany G, Li Y, Wang Y, Ball HL, et al. Yersinia YopJ acetylates and inhibits kinase activation by blocking phosphorylation. Science. 2006;312:1211–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
