

Characterization of granulations of calcium and apatite in serum as pleomorphic mineralo-protein complexes and as precursors of putative nanobacteria
- PMID: 19412552
- PMCID: PMC2673041
- DOI: 10.1371/journal.pone.0005421
Characterization of granulations of calcium and apatite in serum as pleomorphic mineralo-protein complexes and as precursors of putative nanobacteria
Abstract
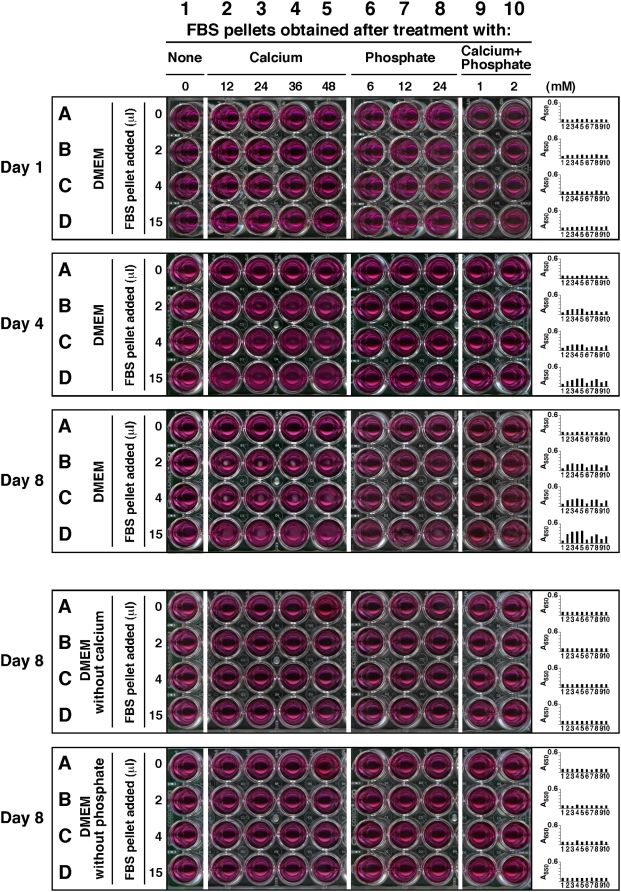
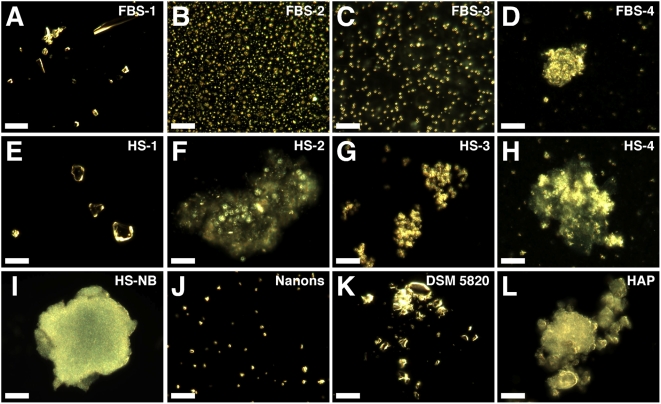
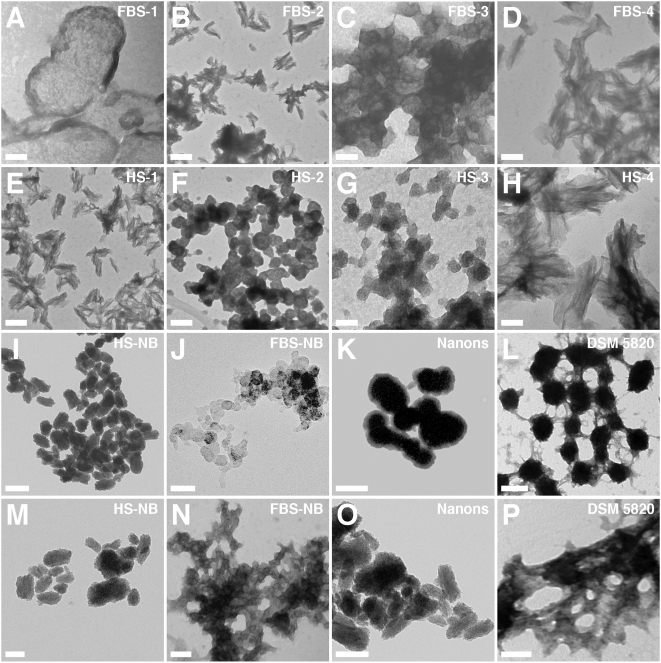
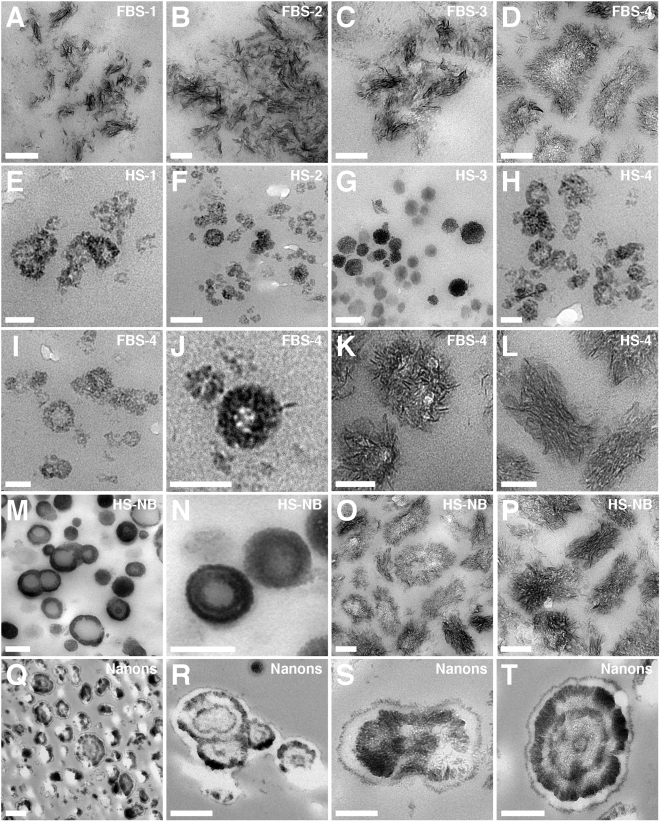
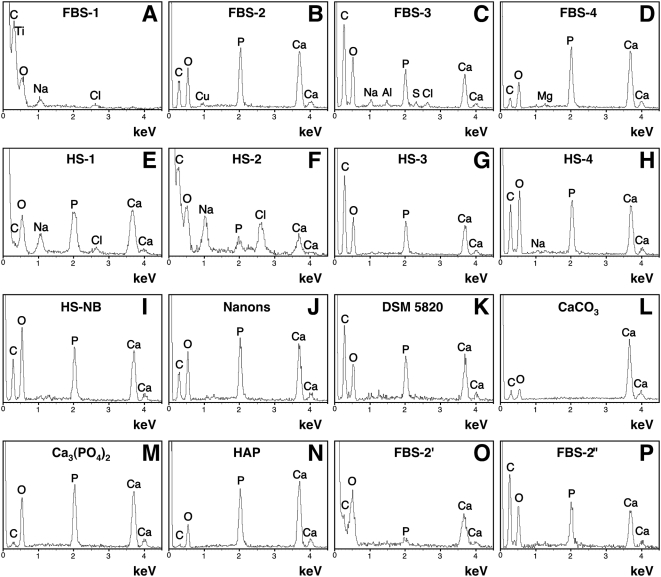
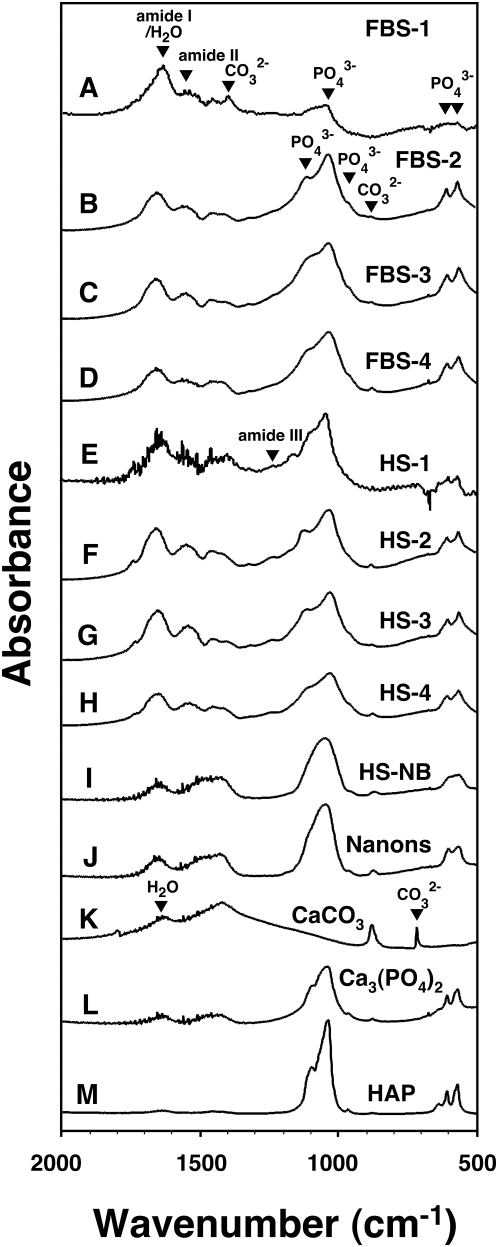
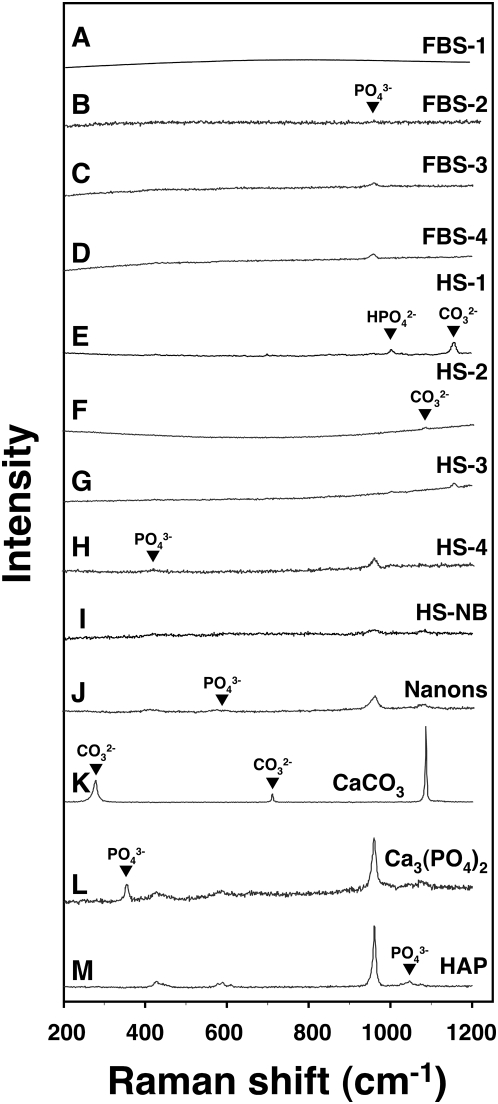
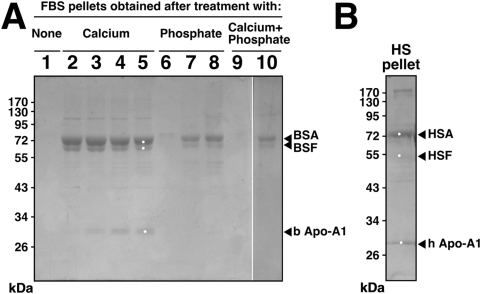
Calcium and apatite granulations are demonstrated here to form in both human and fetal bovine serum in response to the simple addition of either calcium or phosphate, or a combination of both. These granulations are shown to represent precipitating complexes of protein and hydroxyapatite (HAP) that display marked pleomorphism, appearing as round, laminated particles, spindles, and films. These same complexes can be found in normal untreated serum, albeit at much lower amounts, and appear to result from the progressive binding of serum proteins with apatite until reaching saturation, upon which the mineralo-protein complexes precipitate. Chemically and morphologically, these complexes are virtually identical to the so-called nanobacteria (NB) implicated in numerous diseases and considered unusual for their small size, pleomorphism, and the presence of HAP. Like NB, serum granulations can seed particles upon transfer to serum-free medium, and their main protein constituents include albumin, complement components 3 and 4A, fetuin-A, and apolipoproteins A1 and B100, as well as other calcium and apatite binding proteins found in the serum. However, these serum mineralo-protein complexes are formed from the direct chemical binding of inorganic and organic phases, bypassing the need for any biological processes, including the long cultivation in cell culture conditions deemed necessary for the demonstration of NB. Thus, these serum granulations may result from physiologically inherent processes that become amplified with calcium phosphate loading or when subjected to culturing in medium. They may be viewed as simple mineralo-protein complexes formed from the deployment of calcification-inhibitory pathways used by the body to cope with excess calcium phosphate so as to prevent unwarranted calcification. Rather than representing novel pathophysiological mechanisms or exotic lifeforms, these results indicate that the entities described earlier as NB most likely originate from calcium and apatite binding factors in the serum, presumably calcification inhibitors, that upon saturation, form seeds for HAP deposition and growth. These calcium granulations are similar to those found in organisms throughout nature and may represent the products of more general calcium regulation pathways involved in the control of calcium storage, retrieval, tissue deposition, and disposal.
Conflict of interest statement
Figures



References
-
- Kajander EO, Kuronen I, Åkerman K, Pelttari A, Çiftçioglu N. Nanobacteria from blood, the smallest culturable autonomously replicating agent on Earth. Proc Soc Photo Opt Instrum Eng. 1997;3111:420–428.
-
- Kajander EO. Nanobacteria: propagating calcifying nanoparticles. Lett Appl Microbiol. 2006;42:549–552. - PubMed
-
- Ciftcioglu N, McKay DS, Mathew G, Kajander EO. Nanobacteria: fact or fiction? Characteristics, detection, and medical importance of novel self-replicating, calcifying nanoparticles. J Investig Med. 2006;54:385–394. - PubMed
-
- Çiftçioglu N, Björklund M, Kuorikoski K, Bergström K, Kajander EO. Nanobacteria: an infectious cause for kidney stone formation. Kidney Int. 1999;56:1893–1898. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
