

Casein kinase 1 delta regulates the pace of the mammalian circadian clock
- PMID: 19414593
- PMCID: PMC2704743
- DOI: 10.1128/MCB.00338-09
Casein kinase 1 delta regulates the pace of the mammalian circadian clock
Abstract
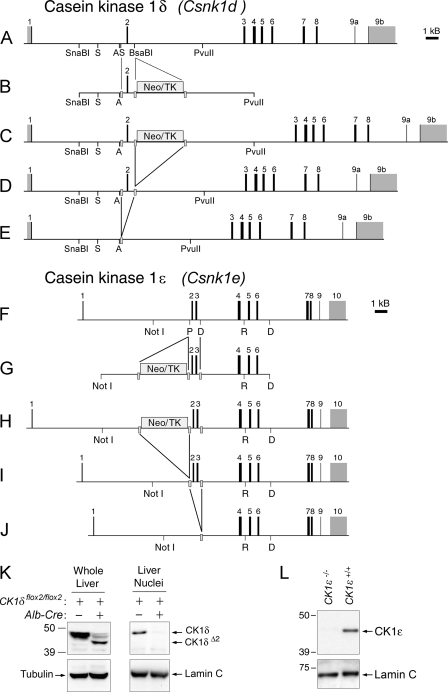
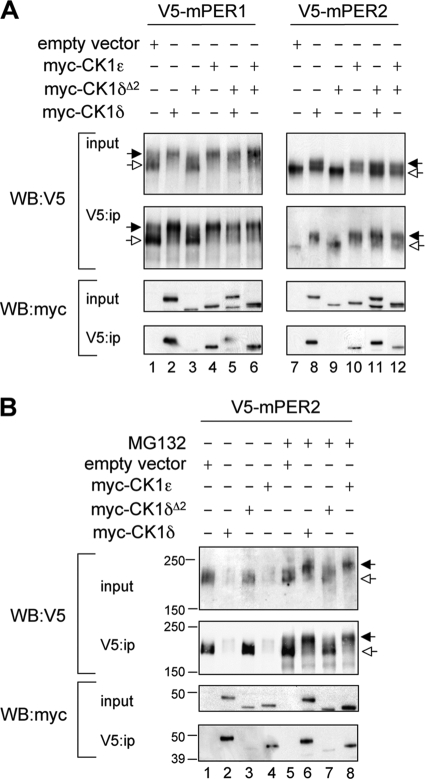
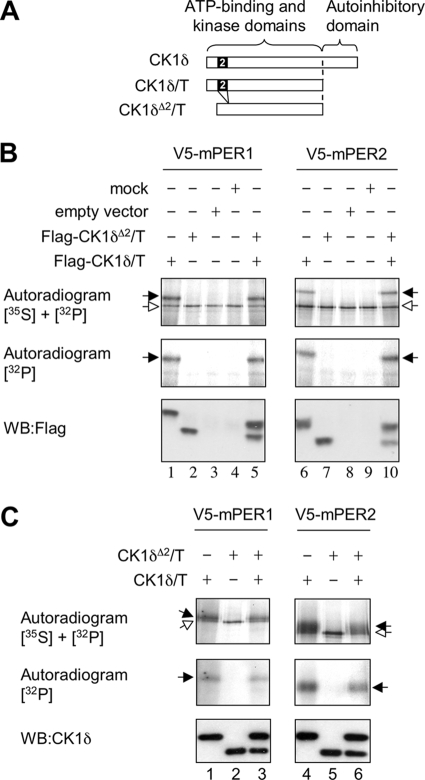
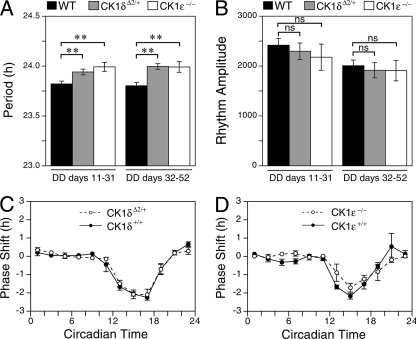
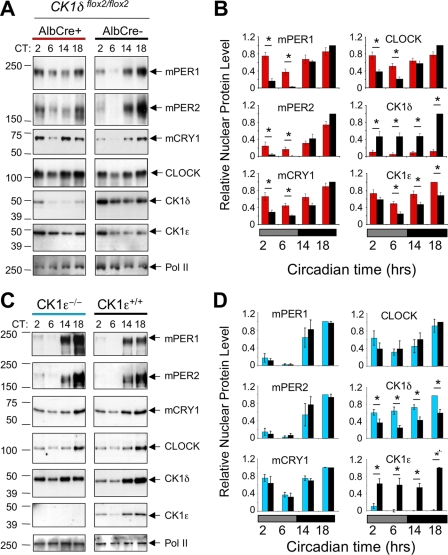
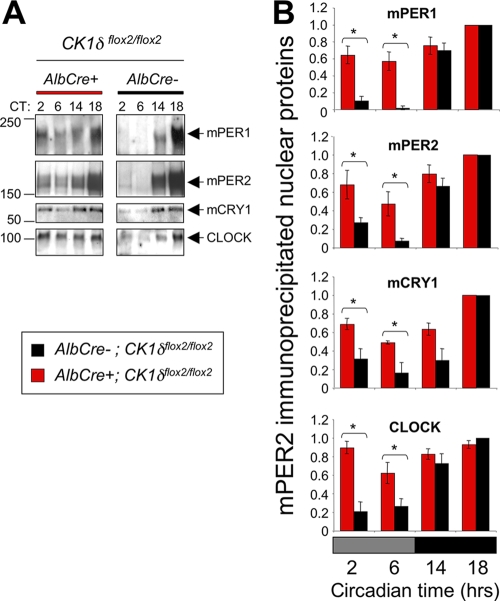
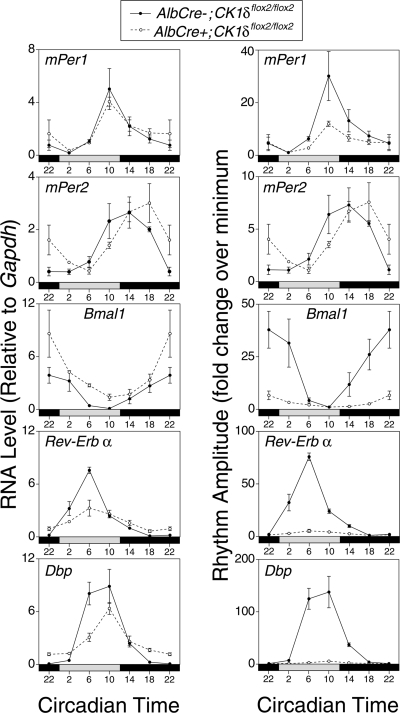
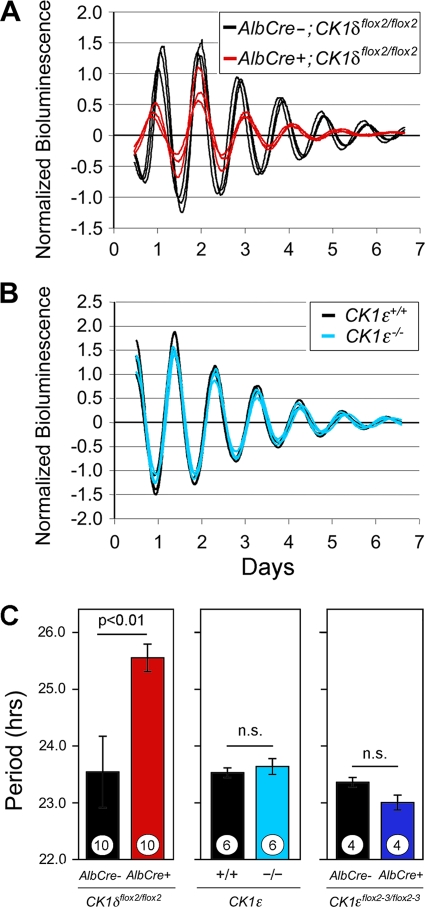
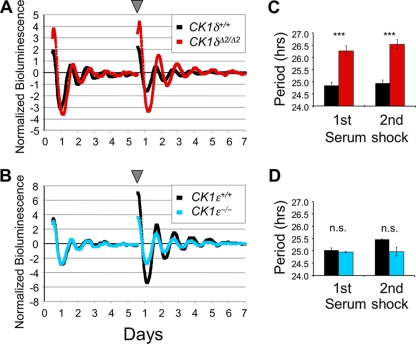
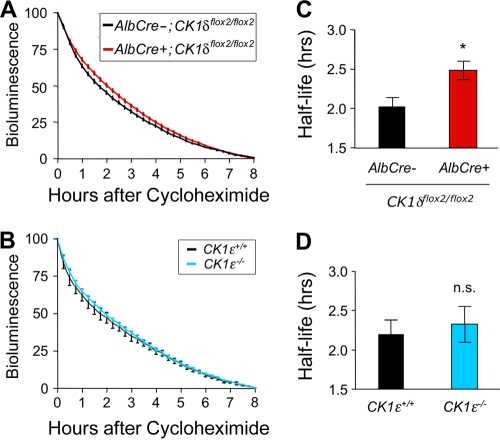
Both casein kinase 1 delta (CK1delta) and epsilon (CK1epsilon) phosphorylate core clock proteins of the mammalian circadian oscillator. To assess the roles of CK1delta and CK1epsilon in the circadian clock mechanism, we generated mice in which the genes encoding these proteins (Csnk1d and Csnk1e, respectively) could be disrupted using the Cre-loxP system. Cre-mediated excision of the floxed exon 2 from Csnk1d led to in-frame splicing and production of a deletion mutant protein (CK1delta(Delta2)). This product is nonfunctional. Mice homozygous for the allele lacking exon 2 die in the perinatal period, so we generated mice with liver-specific disruption of CK1delta. In livers from these mice, daytime levels of nuclear PER proteins, and PER-CRY-CLOCK complexes were elevated. In vitro, the half-life of PER2 was increased by approximately 20%, and the period of PER2::luciferase bioluminescence rhythms was 2 h longer than in controls. Fibroblast cultures from CK1delta-deficient embryos also had long-period rhythms. In contrast, disruption of the gene encoding CK1epsilon did not alter these circadian endpoints. These results reveal important functional differences between CK1delta and CK1epsilon: CK1delta plays an unexpectedly important role in maintaining the 24-h circadian cycle length.
Figures
References
-
- Agostino, P. V., S. A. Plano, and D. A. Golombek. 2008. 2008. Circadian and pharmacological regulation of casein kinase I in the hamster suprachiasmatic nucleus. J. Genet. 87467-471. - PubMed
-
- Badura, L., T. Swanson, W. Adamowicz, J. Adams, J. Cianfrogna, K. Fisher, J. Holland, R. Kleiman, F. Nelson, L. Reynolds, K. St Germain, E. Schaeffer, B. Tate, and J. Sprouse. 2007. An inhibitor of casein kinase I epsilon induces phase delays in circadian rhythms under free-running and entrained conditions. J. Pharmacol. Exp. Ther. 322730-738. - PubMed
-
- Balsalobre, A., F. Damiola, and U. Schibler. 1998. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell 93929-937. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases