

Comparative assessment of large-scale proteomic studies of apoptotic proteolysis
- PMID: 19415908
- PMCID: PMC3150830
- DOI: 10.1021/cb900082q
Comparative assessment of large-scale proteomic studies of apoptotic proteolysis
Abstract
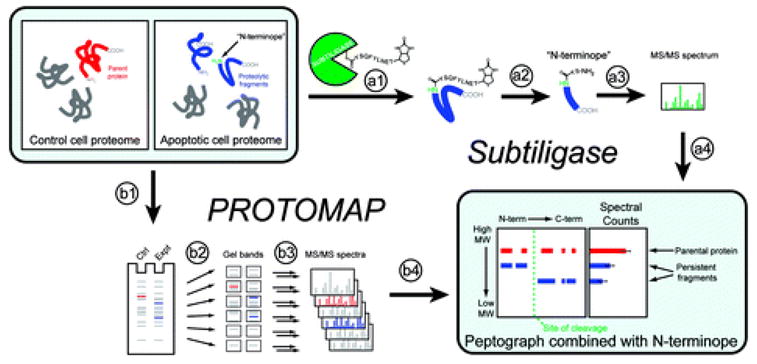
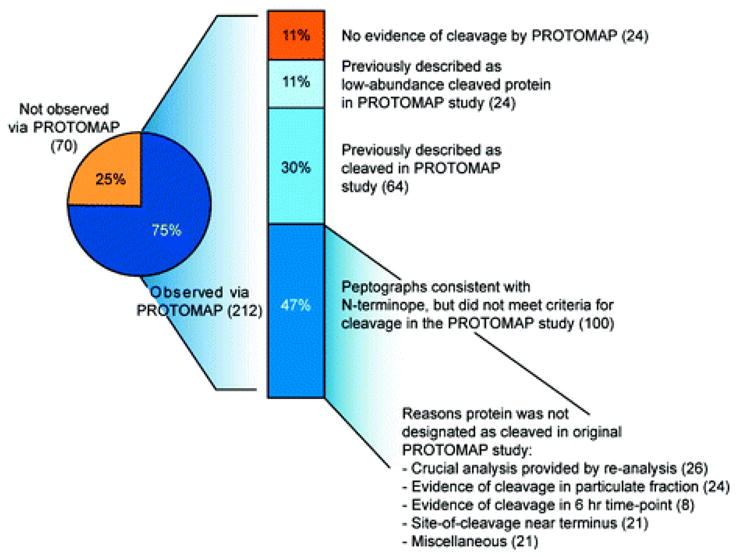
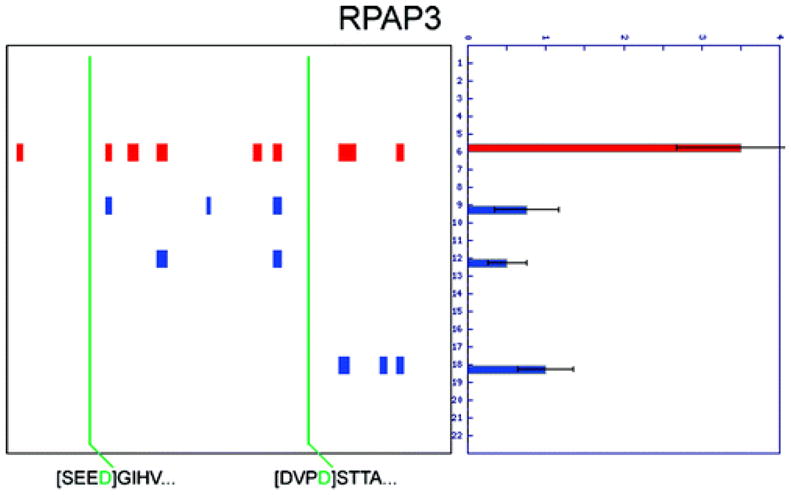
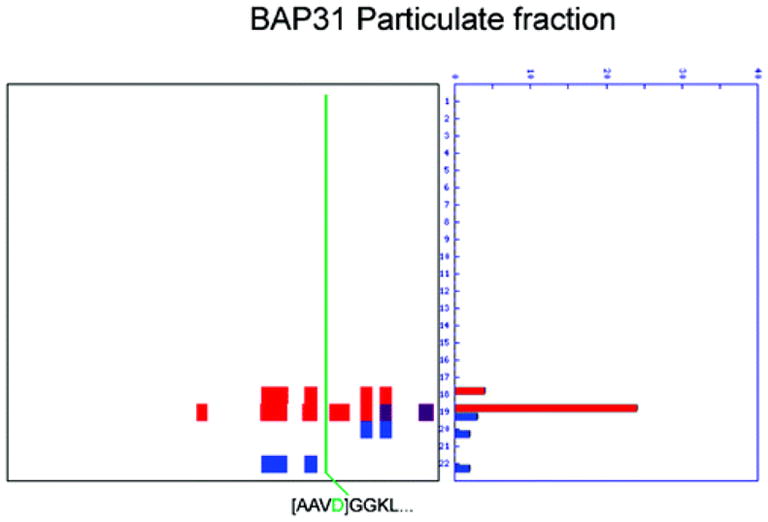
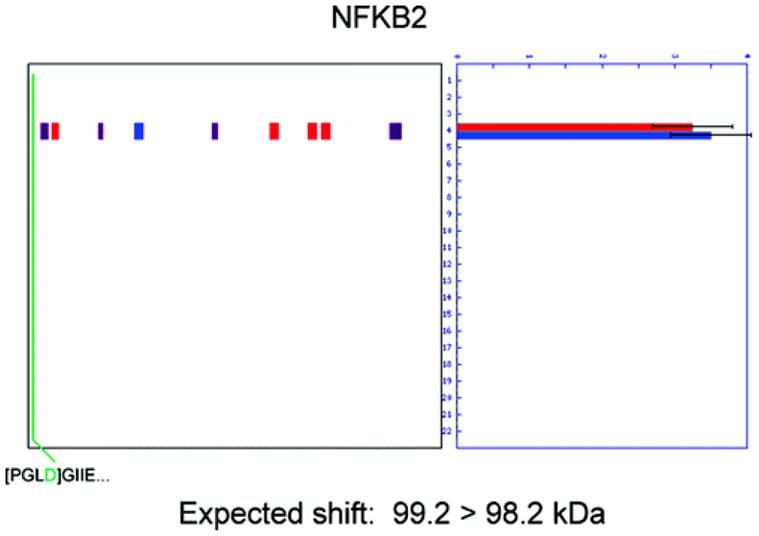
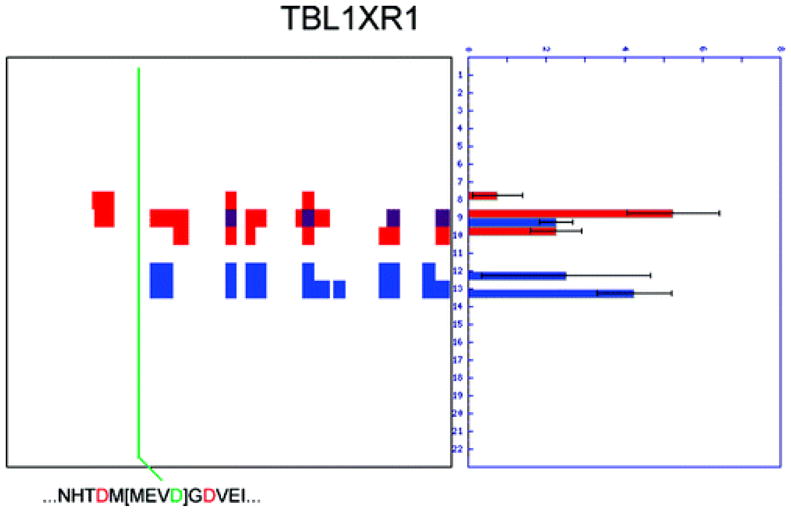
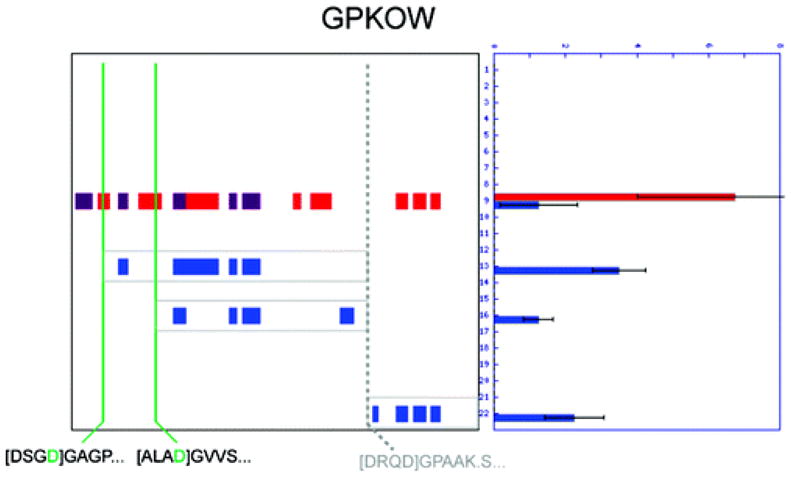
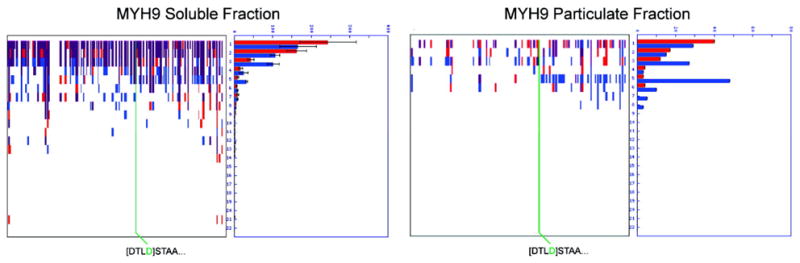
Two proteomic methods were recently introduced to globally map proteolytic cleavage events in biological systems, one that characterized proteolyzed proteins by differential gel migration (PROTOMAP) and the other by enzymatic tagging and enrichment of the nascent N-terminal peptides generated by proteolysis (Subtiligase). Both technologies were applied to apoptosis, and each uncovered hundreds of novel proteolytic events. An initial survey, however, revealed only minimal overlap in the two data sets. In this article, we perform an in-depth comparative analysis of the PROTOMAP and Subtiligase results that assimilates the complementary information acquired by each method. This analysis uncovered substantial agreement between the PROTOMAP and Subtiligase data sets, which in integrated form yield a highly enriched portrait of the proteome-wide impact of proteolysis in apoptosis. We discuss the respective strengths of each proteomic method and the potential for these technologies to expand the scope and sensitivity of large-scale studies of proteolysis in biological systems.
Figures
References
-
- Danial NN, Korsmeyer SJ. Cell death: critical control points. Cell. 2004;116:205–219. - PubMed
-
- Van Damme P, Martens L, Van Damme J, Hugelier K, Staes A, Vandekerckhove J, Gevaert K. Caspase-specific and nonspecific in vivo protein processing during Fas-induced apoptosis. Nat Methods. 2005;2:771–777. - PubMed
-
- Lee AY, Park BC, Jang M, Cho S, Lee DH, Lee SC, Myung PK, Park SG. Identification of caspase-3 degradome by two-dimensional gel electrophoresis and matrix-assisted laser desorption/ionization-time of flight analysis. Proteomics. 2004;4:3429–3436. - PubMed
-
- Anke Rickers EB, Markus Y Mapara, Albrecht Otto, Bernd Drken, Kurt Bommert. Inhibition of CPP32 blocks surface IgM-mediated apoptosis and D4-GDI cleavage in human BL60 Burkitt lymphoma cells. Eur J Immunol. 1998;28:296–304. - PubMed
-
- Brockstedt E, Rickers A, Kostka S, Laubersheimer A, Dorken B, Wittmann-Liebold B, Bommert K, Otto A. Identification of apoptosis-associated proteins in a human Burkitt lymphoma cell line. Cleavage of heterogeneous nuclear ribonucleoprotein A1 by caspase 3. J Biol Chem. 1998;273:28057–28064. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
