

Making the cut: central roles of intramembrane proteolysis in pathogenic microorganisms
- PMID: 19421188
- PMCID: PMC2818034
- DOI: 10.1038/nrmicro2130
Making the cut: central roles of intramembrane proteolysis in pathogenic microorganisms
Abstract
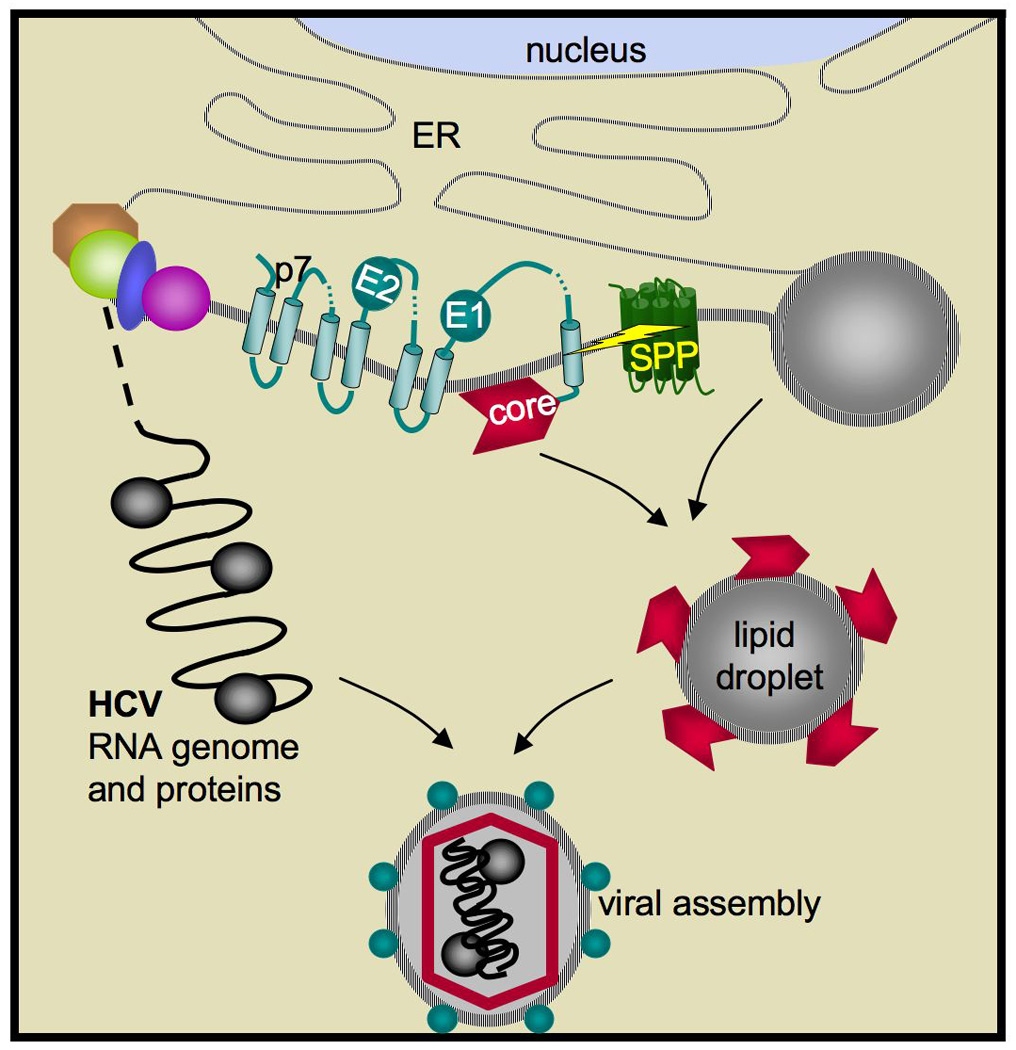
Proteolysis in cellular membranes to liberate effector domains from their transmembrane anchors is a well-studied regulatory mechanism in animal biology and disease. By contrast, the function of intramembrane proteases in unicellular organisms has received little attention. Recent progress has now established that intramembrane proteases execute pivotal roles in a range of pathogens, from regulating Mycobacterium tuberculosis envelope composition, cholera toxin production, bacterial adherence and conjugation, to malaria parasite invasion, fungal virulence, immune evasion by parasitic amoebae and hepatitis C virus assembly. These advances raise the exciting possibility that intramembrane proteases may serve as targets for combating a wide range of infectious diseases. This Review focuses on summarizing the advances, evaluating the limitations and highlighting the promise of this newly emerging field.
Figures





Similar articles
-
Discovery of Cellular Roles of Intramembrane Proteases.ACS Chem Biol. 2019 Nov 15;14(11):2372-2388. doi: 10.1021/acschembio.9b00404. Epub 2019 Jul 24. ACS Chem Biol. 2019. PMID: 31287658 Review.
-
Reconstitution of intramembrane proteolysis in vitro reveals that pure rhomboid is sufficient for catalysis and specificity.Proc Natl Acad Sci U S A. 2005 Feb 8;102(6):1883-8. doi: 10.1073/pnas.0408306102. Epub 2005 Jan 31. Proc Natl Acad Sci U S A. 2005. PMID: 15684070 Free PMC article.
-
Intramembrane proteolysis in regulated protein trafficking.Traffic. 2011 Sep;12(9):1109-18. doi: 10.1111/j.1600-0854.2011.01219.x. Epub 2011 Jun 13. Traffic. 2011. PMID: 21585636 Review.
-
Understanding intramembrane proteolysis: from protein dynamics to reaction kinetics.Trends Biochem Sci. 2015 Jun;40(6):318-27. doi: 10.1016/j.tibs.2015.04.001. Epub 2015 May 1. Trends Biochem Sci. 2015. PMID: 25941170 Review.
-
Intramembrane proteolysis at a glance: from signalling to protein degradation.J Cell Sci. 2019 Aug 15;132(16):jcs217745. doi: 10.1242/jcs.217745. J Cell Sci. 2019. PMID: 31416853 Review.
Cited by
-
Membrane immersion allows rhomboid proteases to achieve specificity by reading transmembrane segment dynamics.Elife. 2012 Nov 13;1:e00173. doi: 10.7554/eLife.00173. Elife. 2012. PMID: 23150798 Free PMC article.
-
Dynamics of the rhomboid-like protein RHBDD2 expression in mouse retina and involvement of its human ortholog in retinitis pigmentosa.J Biol Chem. 2013 Apr 5;288(14):9742-9754. doi: 10.1074/jbc.M112.419960. Epub 2013 Feb 5. J Biol Chem. 2013. PMID: 23386608 Free PMC article.
-
Unraveling the Complexity of the Rhomboid Serine Protease 4 Family of Babesia bovis Using Bioinformatics and Experimental Studies.Pathogens. 2022 Mar 12;11(3):344. doi: 10.3390/pathogens11030344. Pathogens. 2022. PMID: 35335668 Free PMC article.
-
New insights into S2P signaling cascades: regulation, variation, and conservation.Protein Sci. 2010 Nov;19(11):2015-30. doi: 10.1002/pro.496. Protein Sci. 2010. PMID: 20836086 Free PMC article. Review.
-
Comparative Degradome Analysis of the Bovine Piroplasmid Pathogens Babesia bovis and Theileria annulata.Pathogens. 2023 Feb 2;12(2):237. doi: 10.3390/pathogens12020237. Pathogens. 2023. PMID: 36839509 Free PMC article.
References
-
- Brown MS, Ye J, Rawson RB, Goldstein JL. Regulated intramembrane proteolysis: a control mechanism conserved from bacteria to humans. Cell. 2000;100:391–398. - PubMed
-
- Wolfe MS, Kopan R. Intramembrane proteolysis: theme and variations. Science. 2004;305:1119–1123. - PubMed
-
- Urban S. Rhomboid proteins: conserved membrane proteases with divergent biological functions. Genes Dev. 2006;20:3054–3068. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
