

Subventricular zone neural progenitors from rapid brain autopsies of elderly subjects with and without neurodegenerative disease
- PMID: 19425077
- PMCID: PMC2757160
- DOI: 10.1002/cne.22040
Subventricular zone neural progenitors from rapid brain autopsies of elderly subjects with and without neurodegenerative disease
Abstract
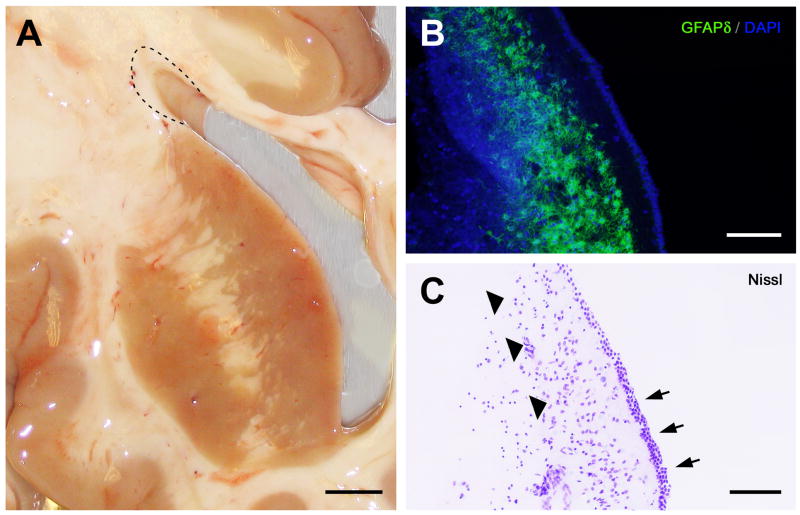
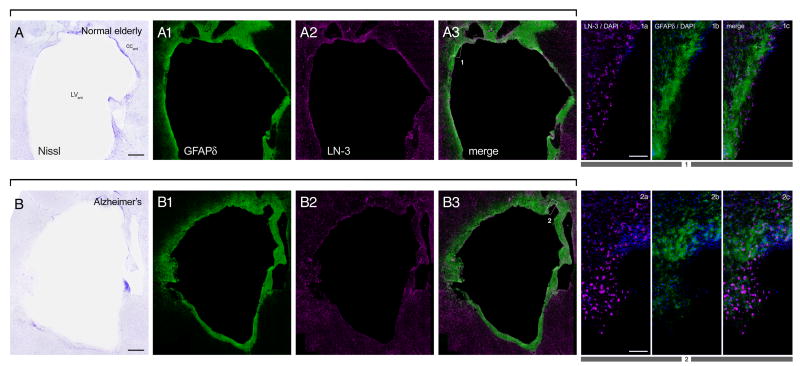
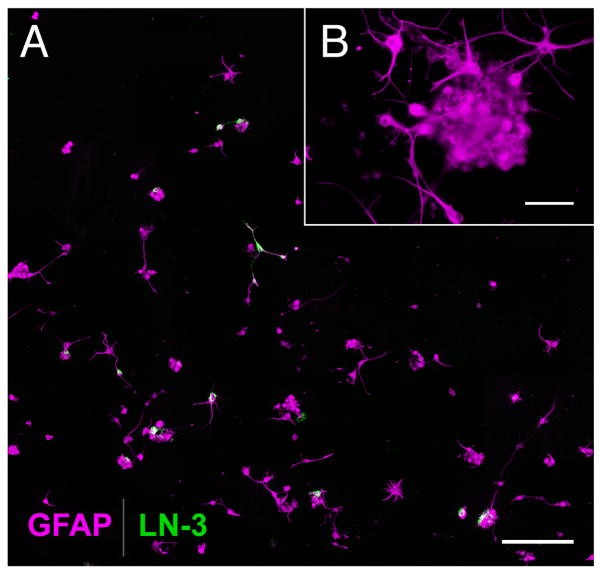
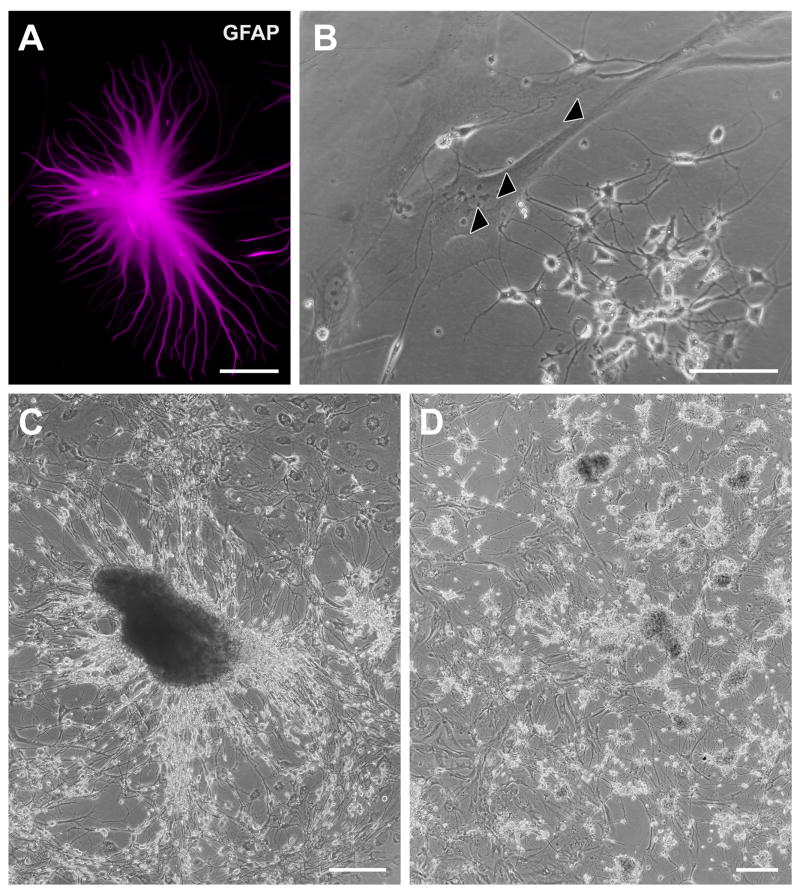
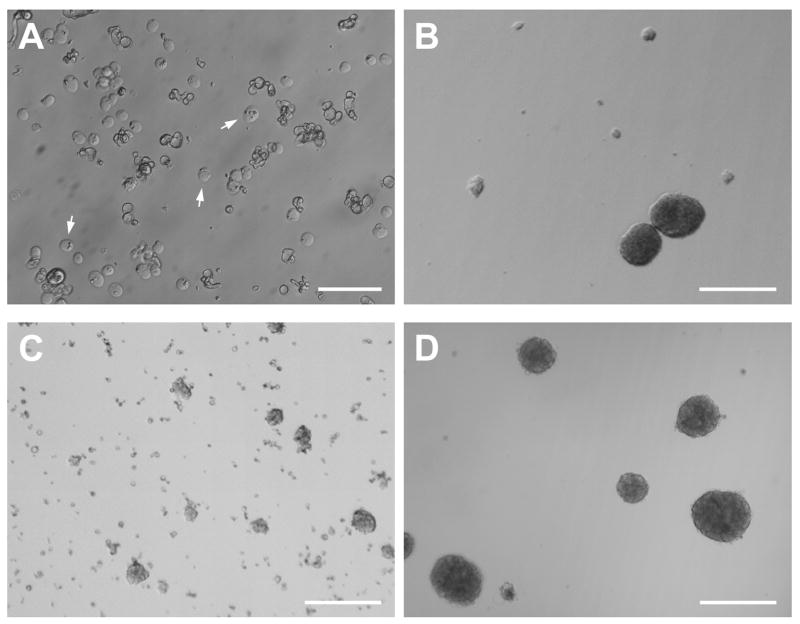
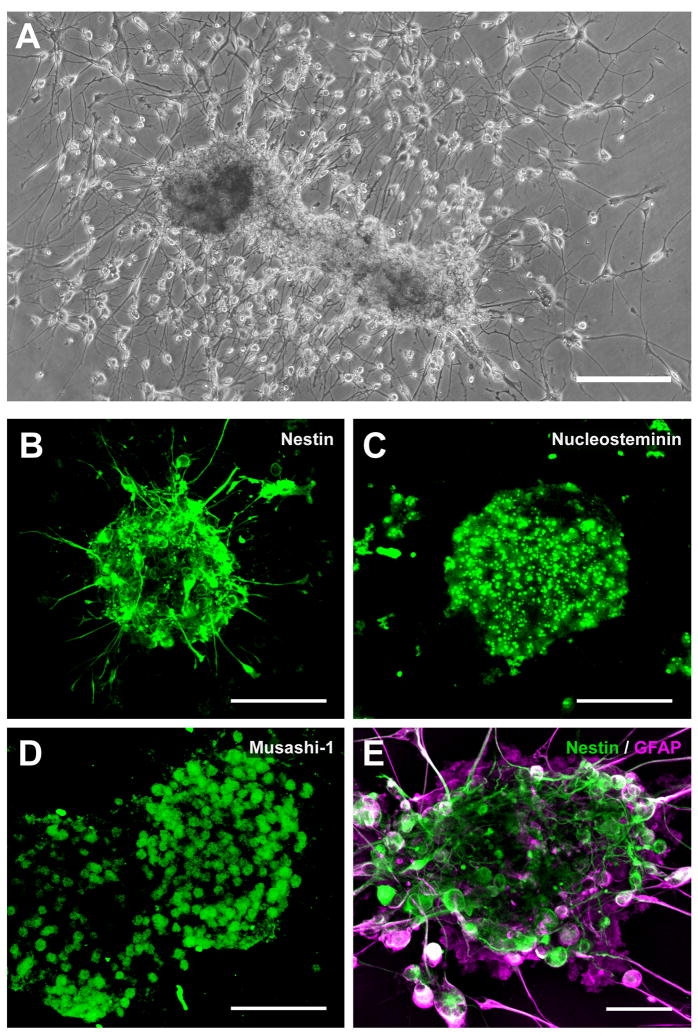
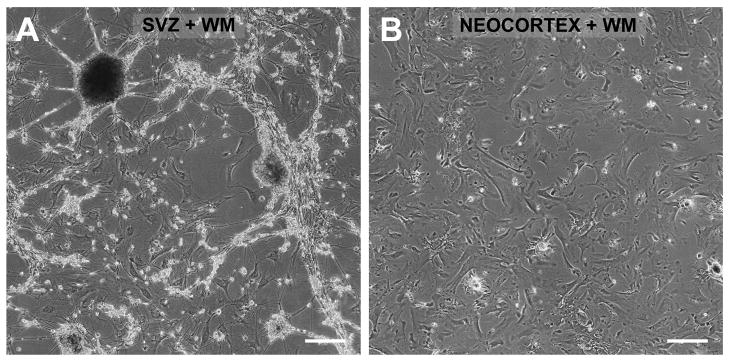
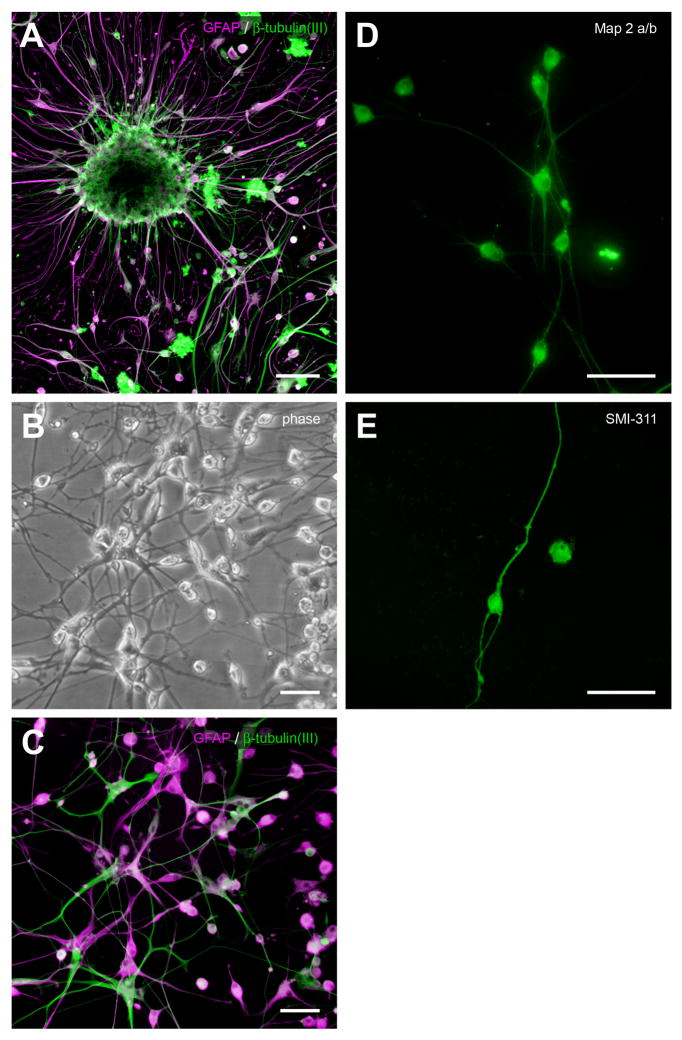
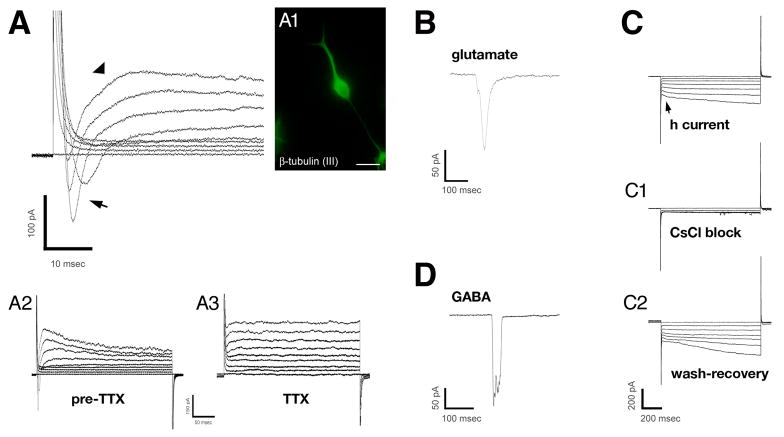
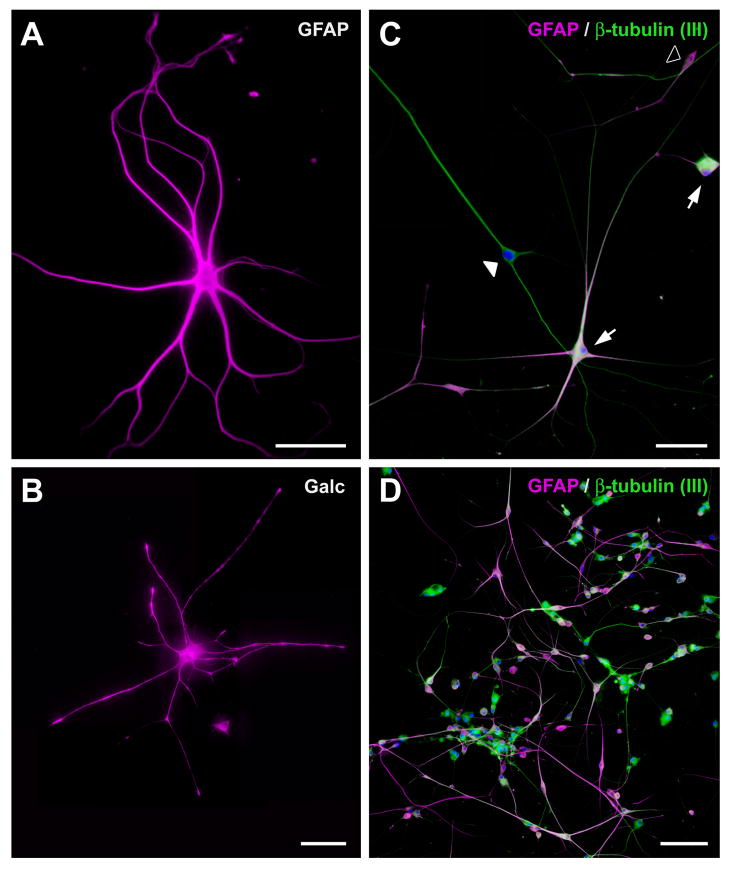
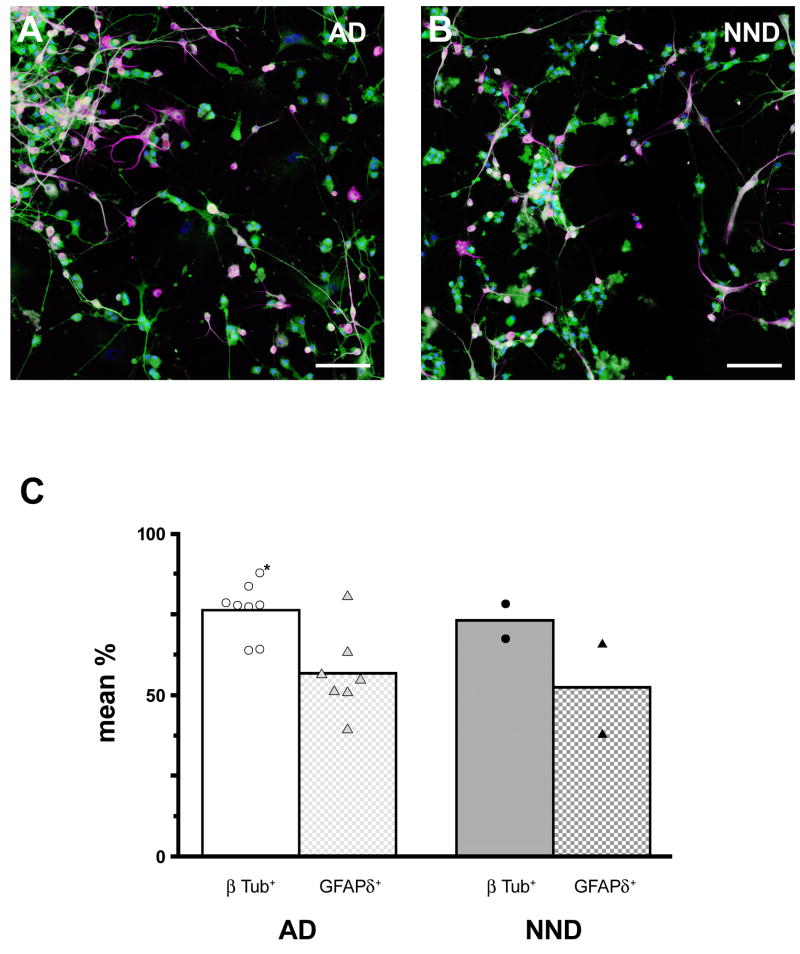
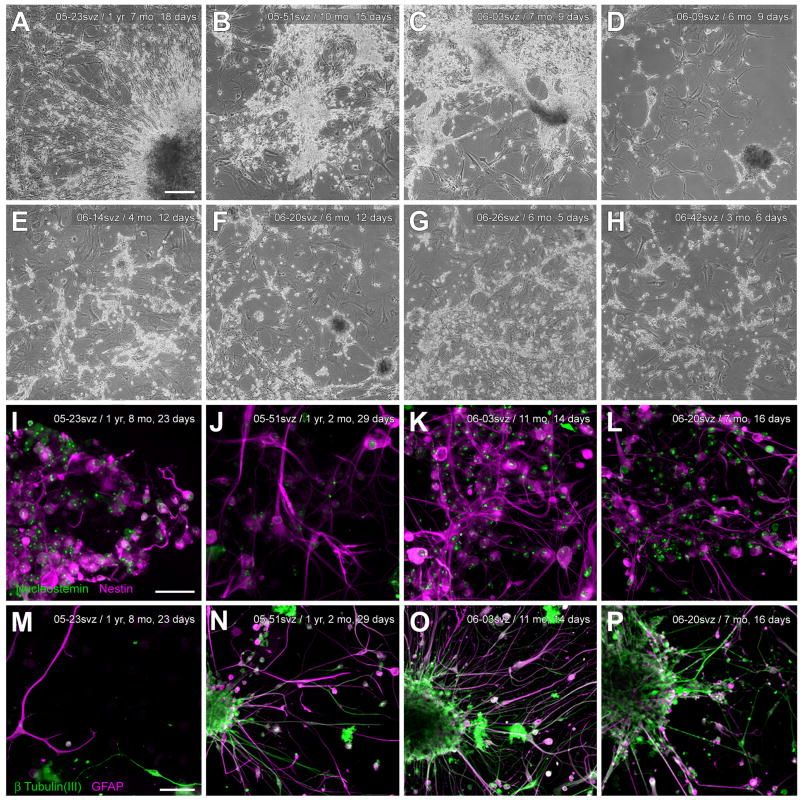
In mice and in young adult humans, the subventricular zone (SVZ) contains multipotent, dividing astrocytes, some of which, when cultured, produce neurospheres that differentiate into neurons and glia. It is unknown whether the SVZ of very old humans has this capacity. Here, we report that neural stem/progenitor cells can also be cultured from rapid autopsy samples of SVZ from elderly human subjects, including patients with age-related neurologic disorders. Histological sections of SVZ from these cases showed a glial fibrillary acidic protein (GFAP)-positive ribbon of astrocytes similar to the astrocyte ribbon in human periventricular white matter biopsies that is reported to be a rich source of neural progenitors. Cultures of the SVZ contained 1) neurospheres with a core of Musashi-1-, nestin-, and nucleostemin-immunopositive cells as well as more differentiated GFAP-positive astrocytes; 2) SMI-311-, MAP2a/b-, and beta-tubulin(III)-positive neurons; and 3) galactocerebroside-positive oligodendrocytes. Neurospheres continued to generate differentiated progeny for months after primary culturing, in some cases nearly 2 years postinitial plating. Patch clamp studies of differentiated SVZ cells expressing neuron-specific antigens revealed voltage-dependent, tetrodotoxin-sensitive, inward Na+ currents and voltage-dependent, delayed, slowly inactivating K+ currents, electrophysiologic characteristics of neurons. A subpopulation of these cells also exhibited responses consistent with the kinetics and pharmacology of the h-current. However, although these cells displayed some aspects of neuronal function, they remained immature, insofar as they did not fire action potentials. These studies suggest that human neural progenitor activity may remain viable throughout much of the life span, even in the face of severe neurodegenerative disease.
Copyright 2009 Wiley-Liss, Inc.
Figures

References
-
- Alafuzoff I, Pikkarainen M, Arzberger T, Thal DR, Al-Sarraj S, Bell J, Bodi I, Budka H, Capetillo-Zarate E, Ferrer I, Gelpi E, Gentleman S, Giaccone G, Kavantzas N, King A, Korkolopoulou P, Kovacs GG, Meyronet D, Monoranu C, Parchi P, Patsouris E, Roggendorf W, Stadelmann C, Streichenberger N, Tagliavini F, Kretzschmar H. Inter-laboratory comparison of neuropathological assessments of beta-amyloid protein: a study of the BrainNet Europe consortium. Acta neuropathologica. 2008;115(5):533–546. - PubMed
-
- Alvarez-Buylla A, Herrera DG, Wichterle H. The subventricular zone: source of neuronal precursors for brain repair. Prog Brain Res. 2000;127:1–11. - PubMed
-
- Banerjee A, Roach MC, Trcka P, Luduena RF. Increased microtubule assembly in bovine brain tubulin lacking the type III isotype of beta-tubulin. J Biol Chem. 1990;265(3):1794–1799. - PubMed
-
- Benjamins JA, Callahan RE, Montgomery IN, Studzinski DM, Dyer CA. Production and characterization of high titer antibodies to galactocerebroside. J Neuroimmunol. 1987;14(3):325–338. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
