

Roles of rel(Spn) in stringent response, global regulation and virulence of serotype 2 Streptococcus pneumoniae D39
- PMID: 19426208
- PMCID: PMC2739083
- DOI: 10.1111/j.1365-2958.2009.06669.x
Roles of rel(Spn) in stringent response, global regulation and virulence of serotype 2 Streptococcus pneumoniae D39
Abstract
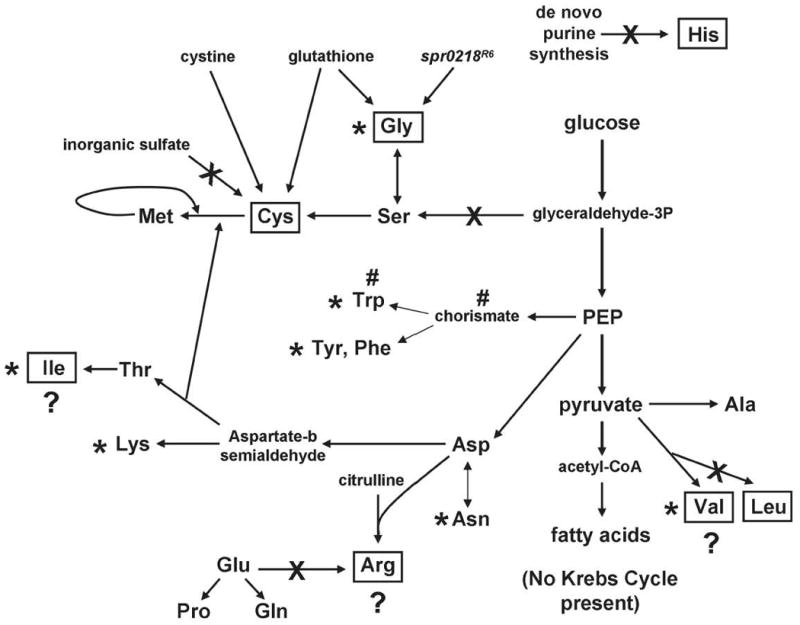
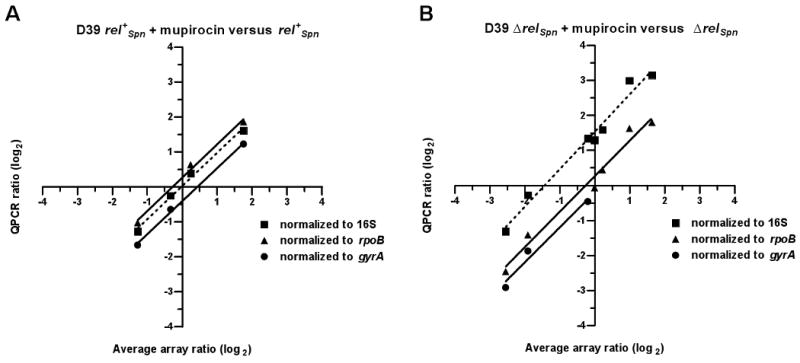
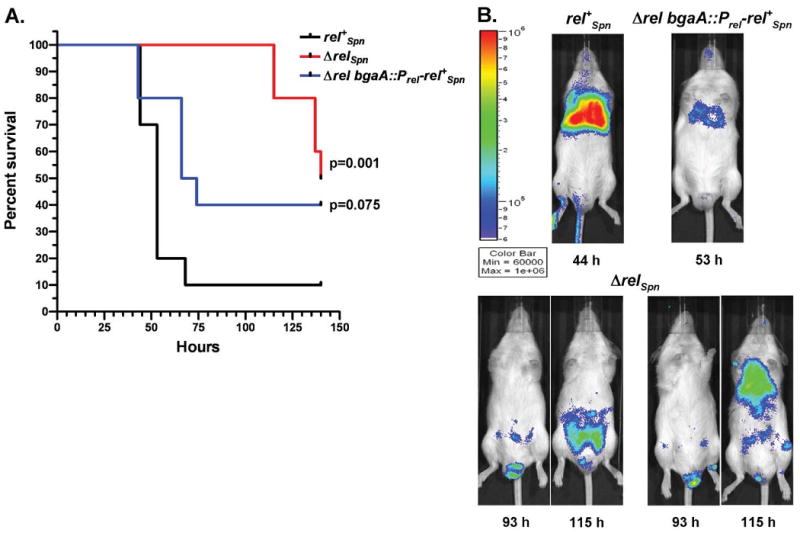
RelA/SpoT homologue (RSH) proteins have (p)ppGpp synthetase and hydrolase activities that mediate major global responses to nutrient limitation and other stresses. RSH proteins are conserved in most bacteria and play diverse roles in bacterial pathogenesis. We report here that the RSH protein of Streptococcus pneumoniae, Rel(Spn), can be deleted and is the primary source of (p)ppGpp synthesis in virulent strain D39 under some conditions. A D39 Deltarel(Spn) mutant grew well in complex medium, but did not grow in chemically defined medium unless supplemented with the metals copper and manganese. Transcriptome analysis of D39 rel(+)(Spn) and Deltarel(Spn) strains treated with mupirocin revealed rel(Spn)-independent (translation stress), rel(Spn)-dependent (stringent response) and Deltarel(Spn)-dependent changes, suggesting that rel(Spn) and (p)ppGpp amount play wide-ranging homeostatic roles in pneumococcal physiology, besides adjusting macromolecular synthesis and transport in response to nutrient availability. Notably, the rel(Spn)-dependent response included significant upregulation of the ply operon encoding pneumolysin toxin, whereas the Deltarel(Spn)-dependent response affected expression linked to the VicRK and CiaRH two-component systems. Finally, a D39 Deltarel(Spn) mutant was severely attenuated and displayed a significantly altered course of disease progression in a mouse model of infection, which was restored to normal by an ectopic copy of rel(+)(Spn).
Figures
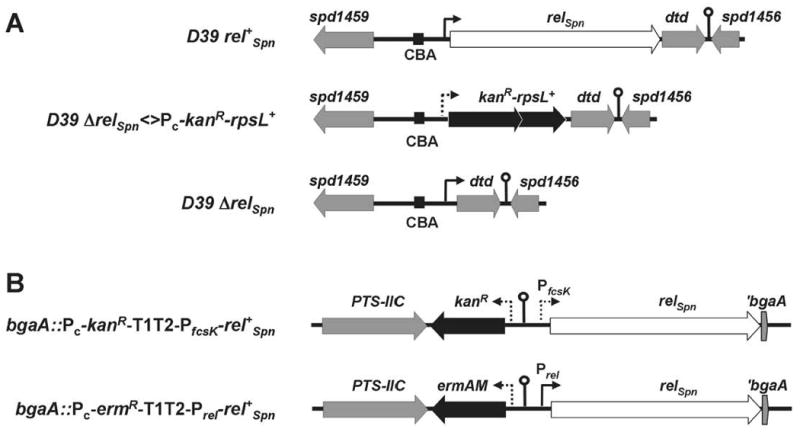
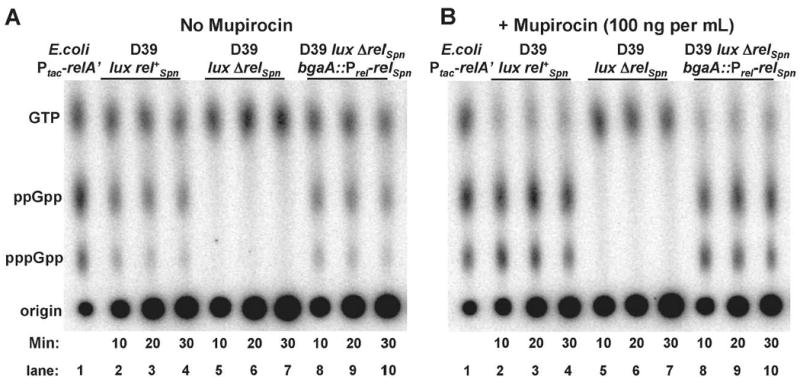
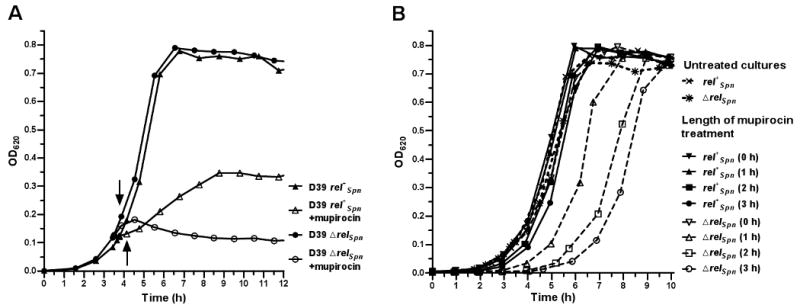
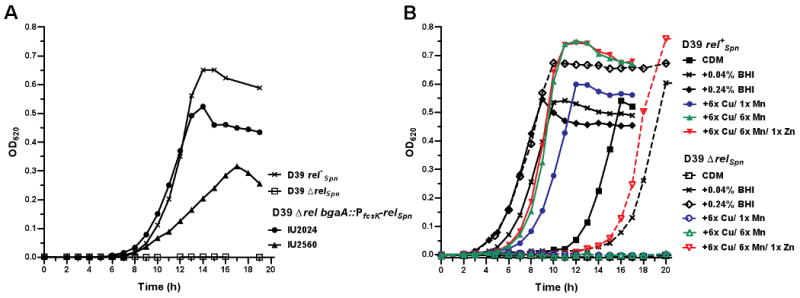
References
-
- Aberg A, Shingler V, Balsalobre C. (p)ppGpp regulates type 1 fimbriation of Escherichia coli by modulating the expression of the site-specific recombinase FimB. Mol Microbiol. 2006;60:1520–1533. - PubMed
-
- Appelbaum PC. Resistance among Streptococcus pneumoniae: Implications for drug selection. Clin Infect Dis. 2002;34:1613–1620. - PubMed
-
- Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B. 1995;57:289–300.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
