

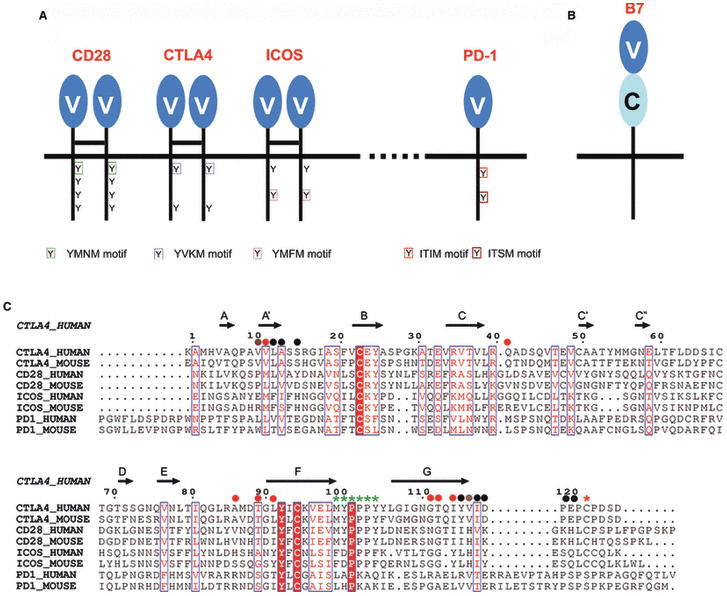
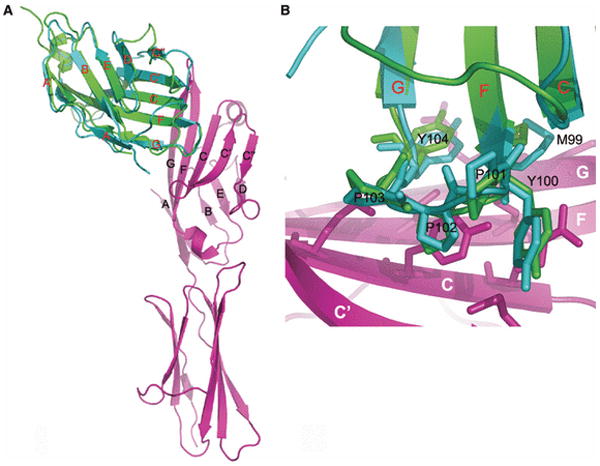
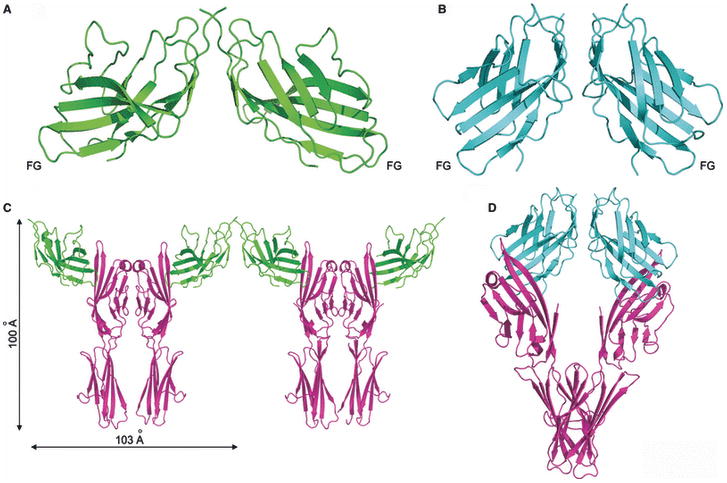
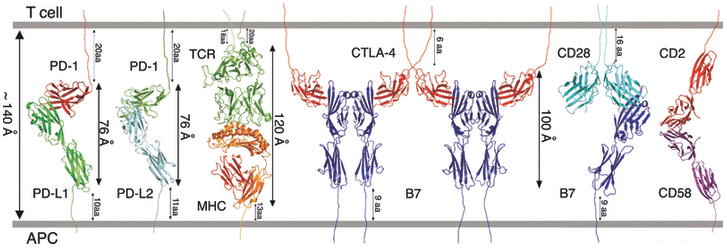
Sequence, structure, function, immunity: structural genomics of costimulation
- PMID: 19426233
- PMCID: PMC3001128
- DOI: 10.1111/j.1600-065X.2009.00778.x
Sequence, structure, function, immunity: structural genomics of costimulation
Abstract
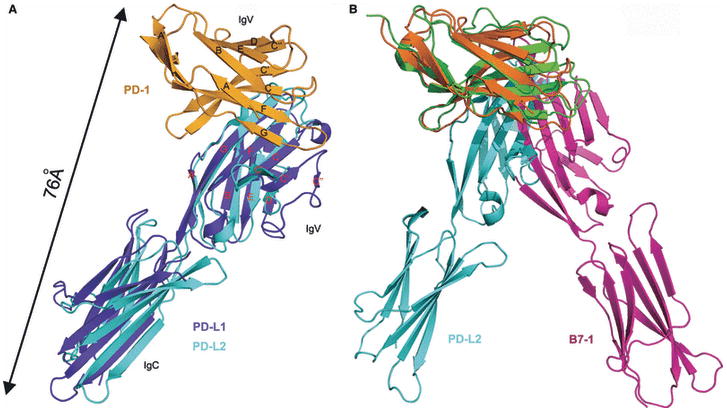
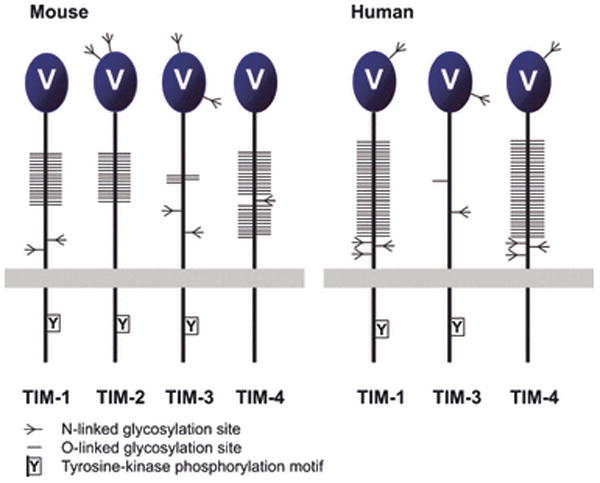
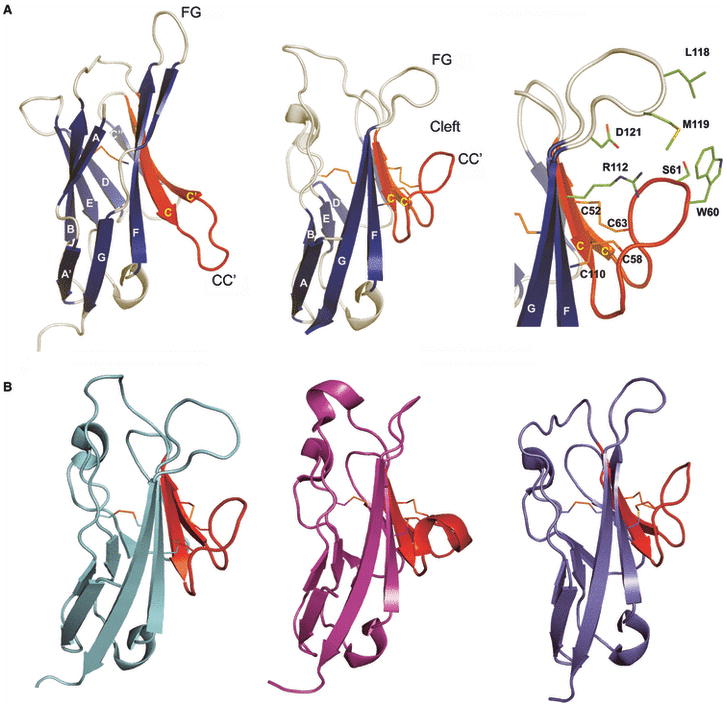
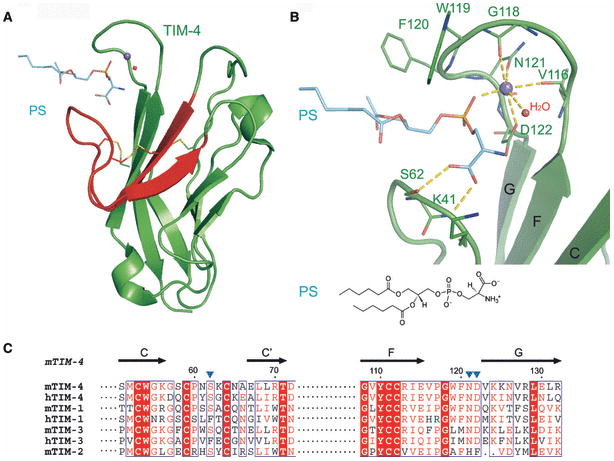
Costimulatory receptors and ligands trigger the signaling pathways that are responsible for modulating the strength, course, and duration of an immune response. High-resolution structures have provided invaluable mechanistic insights by defining the chemical and physical features underlying costimulatory receptor:ligand specificity, affinity, oligomeric state, and valency. Furthermore, these structures revealed general architectural features that are important for the integration of these interactions and their associated signaling pathways into overall cellular physiology. Recent technological advances in structural biology promise unprecedented opportunities for furthering our understanding of the structural features and mechanisms that govern costimulation. In this review, we highlight unique insights that have been revealed by structures of costimulatory molecules from the immunoglobulin and tumor necrosis factor superfamilies and describe a vision for future structural and mechanistic analysis of costimulation. This vision includes simple strategies for the selection of candidate molecules for structure determination and highlights the critical role of structure in the design of mutant costimulatory molecules for the generation of in vivo structure-function correlations in a mammalian model system. This integrated 'atoms-to-animals' paradigm provides a comprehensive approach for defining atomic and molecular mechanisms.
Figures
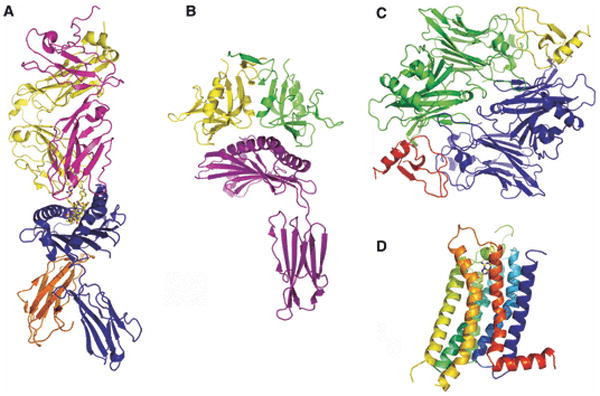
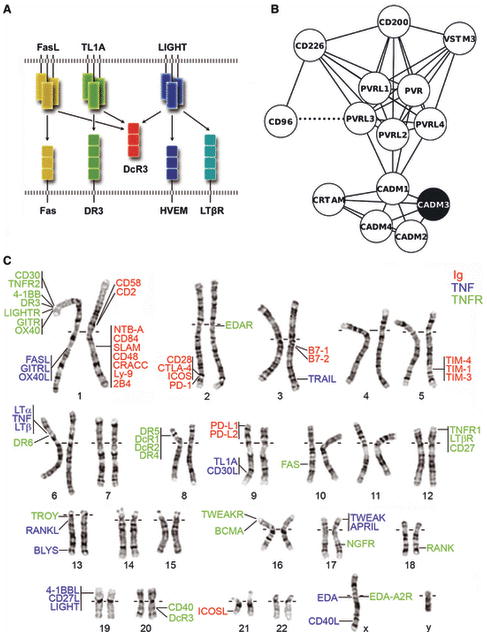
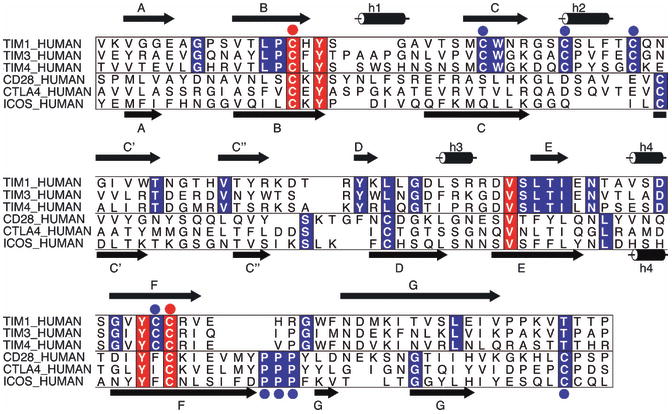
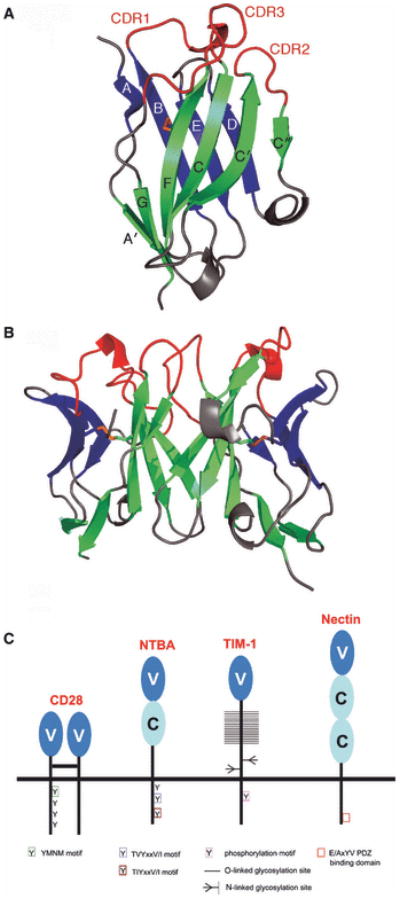
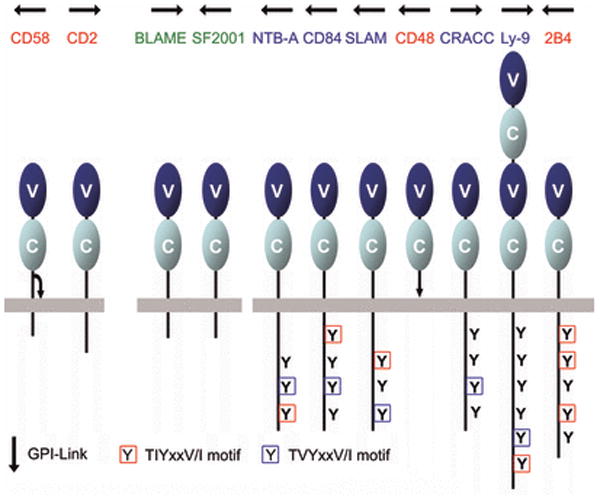
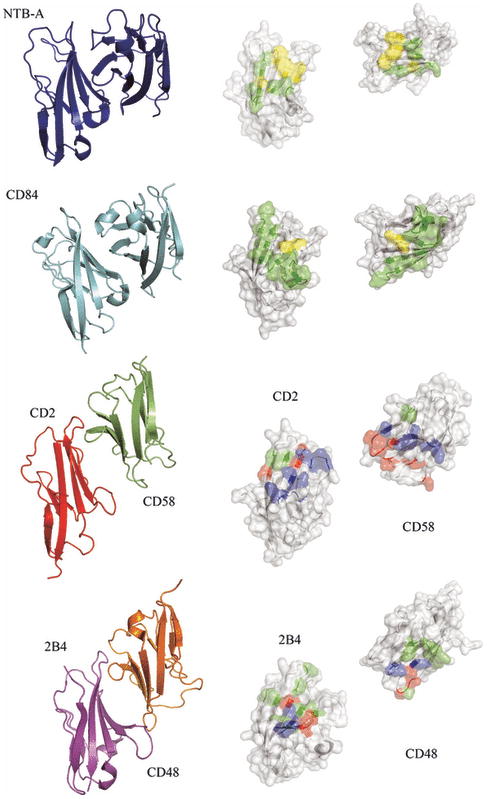
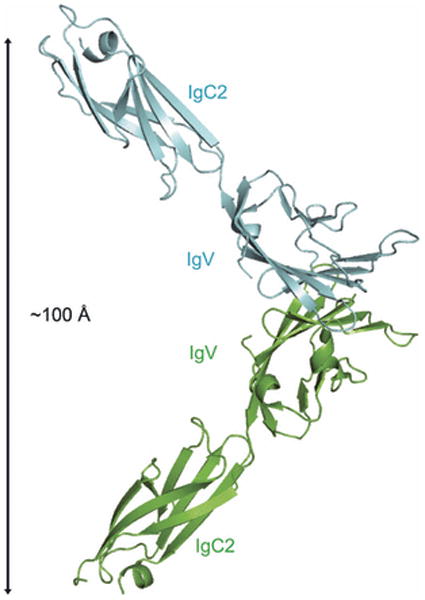
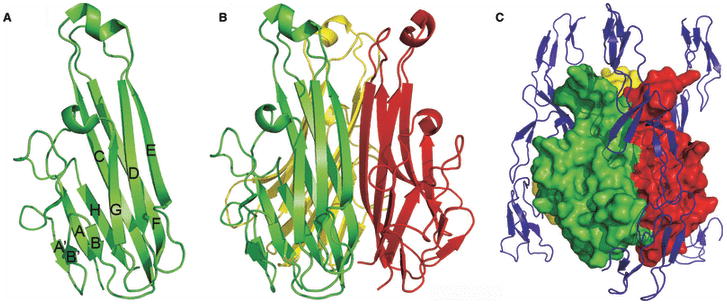
References
-
- Roughton FJW, Kendrew JC. A symposium based on a conference held at Cambridge in June 1948 in memory of Sir Joseph Barcroft. Presented at A symposium based on a conference held at Cambridge in June 1948 in memory of Sir Joseph Barcroft; Cambridge. 1948.
-
- Perutz MF, Muirhead H, Cox JM, Goaman LC. Three-dimensional Fourier synthesis of horse oxyhaemoglobin at 2.8 A resolution: the atomic model. Nature. 1968;219:131–139. - PubMed
-
- Page R, Deacon AM, Lesley SA, Stevens RC. Shotgun crystallization strategy for structural genomics II: crystallization conditions that produce high resolution structures for T. maritima proteins. J Struct Funct Genomics. 2005;6:209–217. - PubMed
-
- Schwartz JC, Zhang X, Nathenson SG, Almo SC. Structural mechanisms of costimulation. Nat Immunol. 2002;3:427–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
