

Synapse formation in developing neural circuits
- PMID: 19427516
- PMCID: PMC7649972
- DOI: 10.1016/S0070-2153(09)01202-2
Synapse formation in developing neural circuits
Abstract
The nervous system consists of hundreds of billions of neurons interconnected into the functional neural networks that underlie behaviors. The capacity of a neuron to innervate and function within a network is mediated via specialized cell junctions known as synapses. Synapses are macromolecular structures that regulate intercellular communication in the nervous system, and are the main gatekeepers of information flow within neural networks. Where and when synapses form determines the connectivity and functionality of neural networks. Therefore, our knowledge of how synapse formation is regulated is critical to our understanding of the nervous system and how it goes awry in neurological disorders. Synapse formation involves pairing of the pre- and postsynaptic partners at a specific neurospatial coordinate. The specificity of synapse formation requires the precise execution of multiple developmental events, including cell fate specification, cell migration, axon guidance, dendritic growth, synaptic target selection, and synaptogenesis (Juttner and Rathjen in Cell. Mol. Life Sci. 62:2811, 2005; Salie et al., in Neuron 45:189, 2005; Waites et al., in Annu. Rev. Neurosci. 28:251, 2005). Remarkably, during the development of the vertebrate nervous system, these developmental processes occur almost simultaneously in billions of neurons, resulting in the formation of trillions of synapses. How this remarkable specificity is orchestrated during development is one of the outstanding questions in the field of neurobiology, and the focus of discussion of this chapter. We center the discussion of this chapter on the early developmental events that orchestrate the process of synaptogenesis prior to activity-dependent mechanisms. We have therefore limited the discussion of important activity-dependent synaptogenic events, which are discussed in other chapters of this book. Moreover, our discussion is biased toward lessons we have learned from invertebrate systems, in particular from C. elegans and Drosophila. We did so to complement the discussions from other chapters in this book, which focus on the important findings that have recently emerged from the vertebrate literature. The chapter begins with a brief history of the field of synaptic biology. This serves as a backdrop to introduce some of the historically outstanding questions of synaptic development that have eluded us during the past century, and which are the focus of this review. We then discuss some general features of synaptic structure as it relates to its function. In particular, we will highlight evolutionarily conserved traits shared by all synaptic structures, and how these features have helped optimize these ancient cellular junctions for interneural communication. We then discuss the regulatory signals that orchestrate the precise assembly of these conserved macromolecular structures. This discussion will be framed in the context of the neurodevelopmental process. Specifically, much of our discussion will focus on how the seemingly disparate developmental processes are intimately linked at a molecular level, and how this relationship might be crucial in the developmental orchestration of circuit assembly. We hope that the discussion of the multifunctional cues that direct circuit development provides a conceptual framework into understanding how, with a limited set of signaling molecules, precise neural wiring can be coordinated between synaptic partners.
Figures
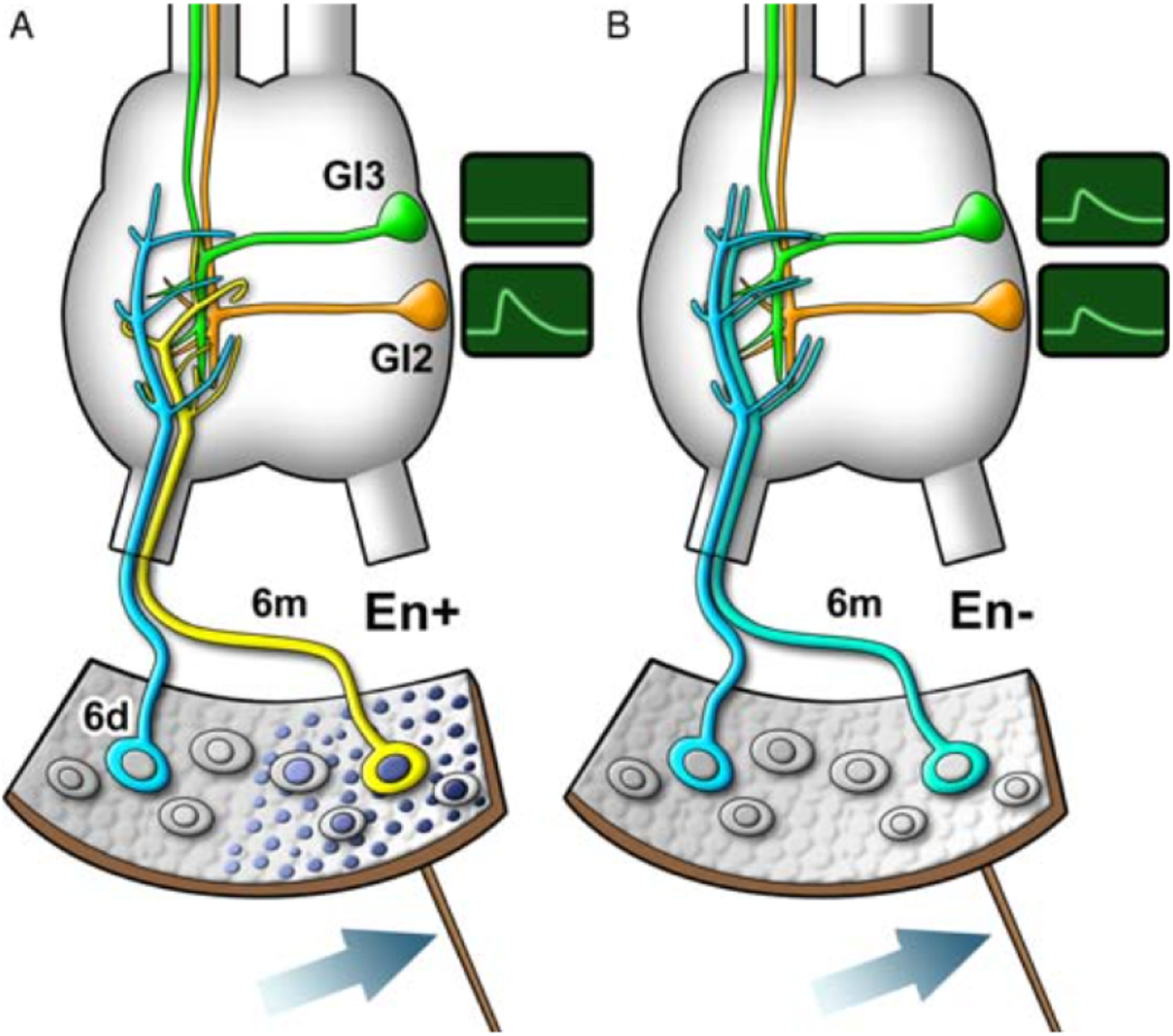


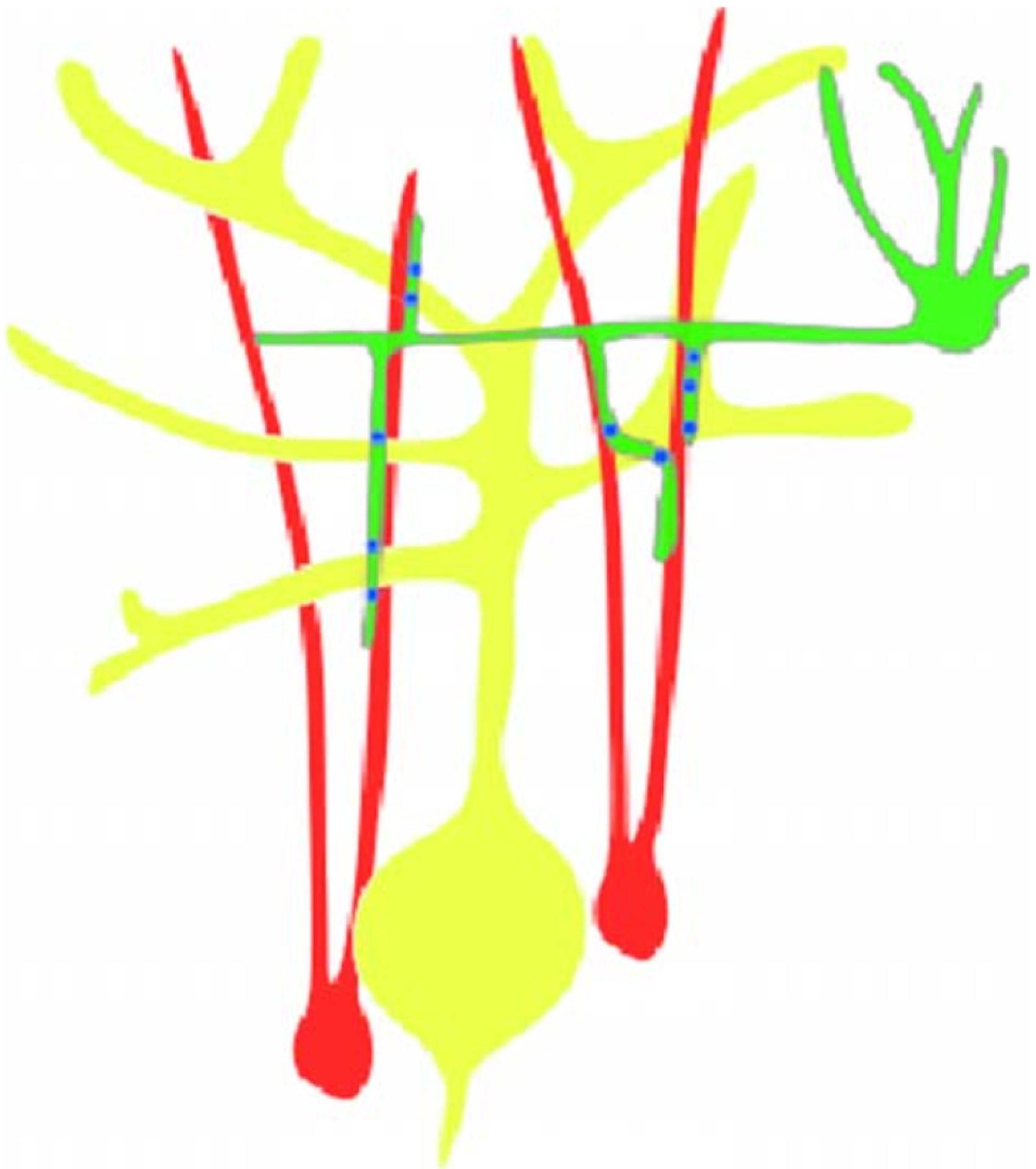
References
-
- Aberle H, Haghighi AP, Fetter RD, McCabe BD, Magalhaes TR, and Goodman CS (2002). Wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron 33, 545–558. - PubMed
-
- Ackley BD, Harrington RJ, Hudson ML, Williams L, Kenyon CJ, Chisholin AD, and Jin Y (2005). The two isoforms of the Caenorhabditis elegans leukocyte-common antigen related receptor tyrosine phosphatase PTP-3 function independently in axon guidance and synapse formation. J. Neurosci 25, 7517–7528. - PMC - PubMed
-
- Akins MR, and Biederer T (2006). Cell-cell interactions in synaptogenesis. Curr. Opin. Neurobiol 16, 83–89. - PubMed
-
- Anderson PA, and Spencer AN (1989). The importance of cnidarian synapses for neurobiology. J. Neurobiol 20, 435–457. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
