

Neural bases of eye and gaze processing: the core of social cognition
- PMID: 19428496
- PMCID: PMC3925117
- DOI: 10.1016/j.neubiorev.2009.02.004
Neural bases of eye and gaze processing: the core of social cognition
Abstract
Eyes and gaze are very important stimuli for human social interactions. Recent studies suggest that impairments in recognizing face identity, facial emotions or in inferring attention and intentions of others could be linked to difficulties in extracting the relevant information from the eye region including gaze direction. In this review, we address the central role of eyes and gaze in social cognition. We start with behavioral data demonstrating the importance of the eye region and the impact of gaze on the most significant aspects of face processing. We review neuropsychological cases and data from various imaging techniques such as fMRI/PET and ERP/MEG, in an attempt to best describe the spatio-temporal networks underlying these processes. The existence of a neuronal eye detector mechanism is discussed as well as the links between eye gaze and social cognition impairments in autism. We suggest impairments in processing eyes and gaze may represent a core deficiency in several other brain pathologies and may be central to abnormal social cognition.
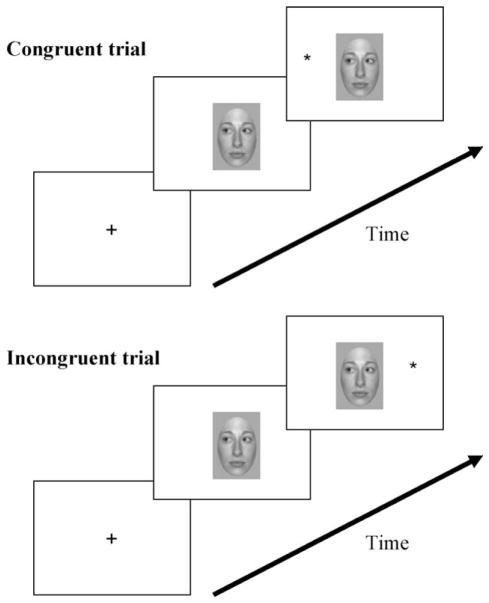
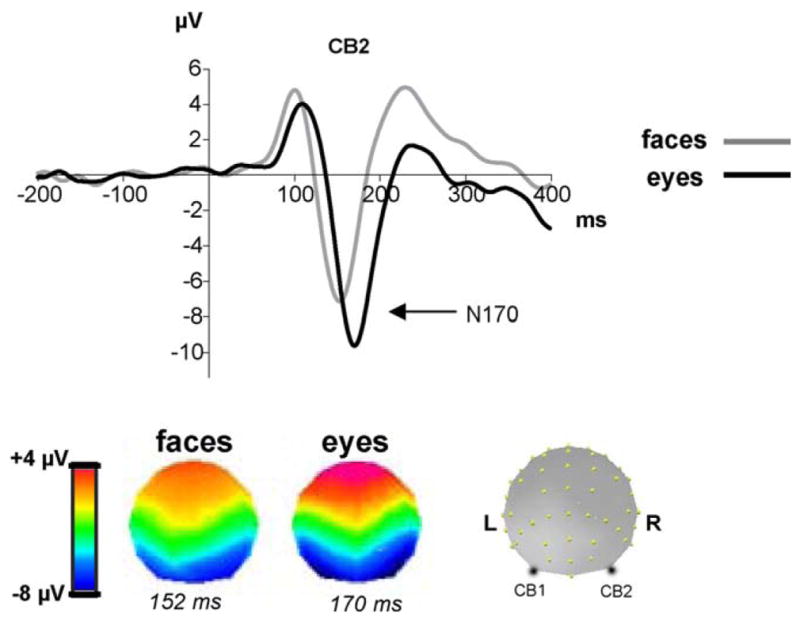
Figures



References
-
- Abell F, Krams M, Ashburner J, Passingham R, Friston K, Frackowiak R, et al. The neuroanatomy of autism: a voxel-based whole brain analysis of structural scans. Neuroreport. 1999;10 (8):1647–1651. - PubMed
-
- Adams RB, Jr, Gordon HL, Baird AA, Ambady N, Kleck RE. Effects of gaze on amygdala sensitivity to anger and fear faces. Science. 2003;300 (5625):1536. - PubMed
-
- Adams RB, Jr, Kleck RE. Perceived gaze direction and the processing of facial displays of emotion. Psychol Sci. 2003;14 (6):644–647. - PubMed
-
- Adams RB, Jr, Kleck RE. Effects of direct and averted gaze on the perception of facially communicated emotion. Emotion. 2005;5 (1):3–11. - PubMed
-
- Adolphs R. Social cognition and the human brain. Trends Cogn Sci. 1999;3 (12):469–479. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
