

Mitochondria and metazoan epigenesis
- PMID: 19429498
- PMCID: PMC2756741
- DOI: 10.1016/j.semcdb.2009.02.002
Mitochondria and metazoan epigenesis
Abstract
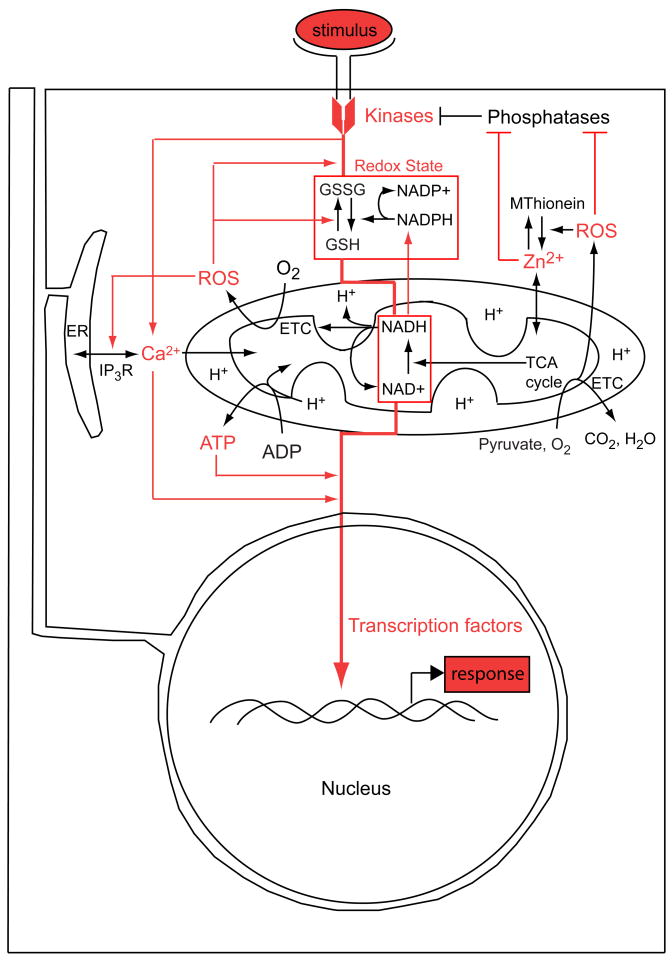
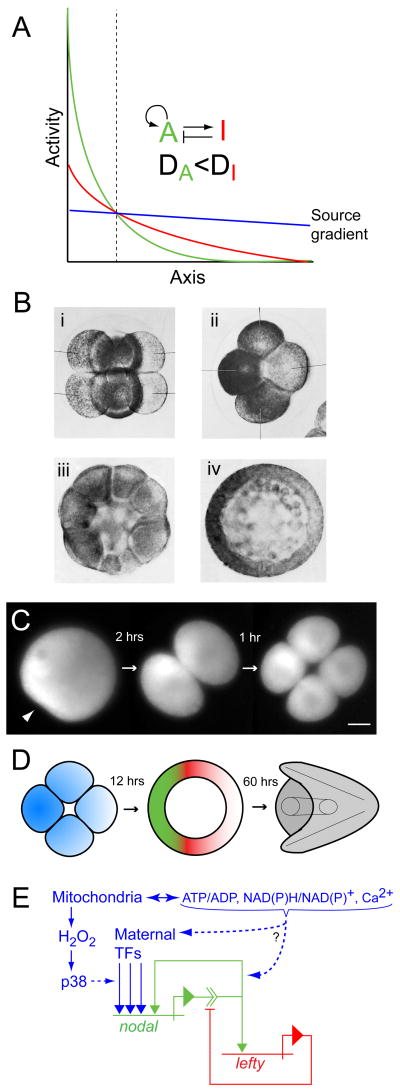
In eukaryotes, mitochondrial activity controls ATP production, calcium dynamics, and redox state, thereby establishing physiological parameters governing the transduction of biochemical signals that regulate nuclear gene expression. However, these activities are commonly assumed to fulfill a 'housekeeping' function: necessary for life, but an epiphenomenon devoid of causal agency in the developmental flow of genetic information. Moreover, it is difficult to perturb mitochondrial function without generally affecting cell viability. For these reasons little is known about the extent of mitochondrial influence on gene activity in early development. Recent discoveries pertaining to the redox regulation of key developmental signaling systems together with the fact that mitochondria are often asymmetrically distributed in animal embryos suggests that they may contribute spatial information underlying differential specification of cell fate. In many cases such asymmetries correlate with localization of genetic determinants (i.e., mRNAs or proteins), particularly in embryos that rely heavily on cell-autonomous means of cell fate specification. In such embryos the localized genetic determinants play a dominant role, and any developmental information contributed by the mitochondria themselves is likely to be less obvious and more difficult to isolate experimentally. Hence, 'regulative' embryos that make more extensive use of conditional cell fate specification are better suited to experimental investigation of mitochondrial impacts on developmental gene regulation. Recent studies of the sea urchin embryo, which is a paradigmatic example of such a system, suggest that anisotropic distribution of mitochondria provides a source gradient of spatial information that directs epigenetic specification of the secondary axis via Nodal-Lefty signaling.
Figures
Similar articles
-
Mitochondria, redox signaling and axis specification in metazoan embryos.Dev Biol. 2007 Aug 15;308(2):266-80. doi: 10.1016/j.ydbio.2007.05.042. Epub 2007 Jun 5. Dev Biol. 2007. PMID: 17586486 Review.
-
Oral-aboral axis specification in the sea urchin embryo II. Mitochondrial distribution and redox state contribute to establishing polarity in Strongylocentrotus purpuratus.Dev Biol. 2004 Sep 1;273(1):160-71. doi: 10.1016/j.ydbio.2004.06.005. Dev Biol. 2004. PMID: 15302605
-
Oral-aboral axis specification in the sea urchin embryo, IV: hypoxia radializes embryos by preventing the initial spatialization of nodal activity.Dev Biol. 2014 Feb 15;386(2):302-7. doi: 10.1016/j.ydbio.2013.12.035. Epub 2013 Dec 31. Dev Biol. 2014. PMID: 24384388 Free PMC article.
-
Nuclear beta-catenin is required to specify vegetal cell fates in the sea urchin embryo.Development. 1999 Jan;126(2):345-57. doi: 10.1242/dev.126.2.345. Development. 1999. PMID: 9847248
-
Maternal control of early patterning in sea urchin embryos.Differentiation. 2020 May-Jun;113:28-37. doi: 10.1016/j.diff.2020.04.001. Epub 2020 Apr 22. Differentiation. 2020. PMID: 32371341 Review.
Cited by
-
Early asymmetric cues triggering the dorsal/ventral gene regulatory network of the sea urchin embryo.Elife. 2014 Dec 2;3:e04664. doi: 10.7554/eLife.04664. Elife. 2014. PMID: 25457050 Free PMC article.
-
Opposing Nodal/Vg1 and BMP signals mediate axial patterning in embryos of the basal chordate amphioxus.Dev Biol. 2010 Aug 1;344(1):377-89. doi: 10.1016/j.ydbio.2010.05.016. Epub 2010 May 19. Dev Biol. 2010. PMID: 20488174 Free PMC article.
-
Adult body weight is programmed by a redox-regulated and energy-dependent process during the pronuclear stage in mouse.PLoS One. 2011;6(12):e29388. doi: 10.1371/journal.pone.0029388. Epub 2011 Dec 28. PLoS One. 2011. PMID: 22216268 Free PMC article.
-
Effects of Caffeic Acid Phenethyl Ester on Embryonic Development Through Regulation of Mitochondria and Endoplasmic Reticulum.Vet Sci. 2024 Dec 6;11(12):625. doi: 10.3390/vetsci11120625. Vet Sci. 2024. PMID: 39728965 Free PMC article.
-
Female and male gamete mitochondria are distinct and complementary in transcription, structure, and genome function.Genome Biol Evol. 2013;5(10):1969-77. doi: 10.1093/gbe/evt147. Genome Biol Evol. 2013. PMID: 24068653 Free PMC article.
References
-
- Coffman JA, Denegre JM. Mitochondria, redox signaling and axis specification in metazoan embryos. Dev Biol. 2007;308:266–80. - PubMed
-
- Child CM. Patterns and Problems of Development. Chicago, IL: University of Chicago Press; 1941.
-
- Covarrubias L, et al. Function of reactive oxygen species during animal development: passive or active? Dev Biol. 2008;320:1–11. - PubMed
-
- Finkel T. Oxidant signals and oxidative stress. Curr Opin Cell Biol. 2003;15:247–54. - PubMed
-
- Connor KM, et al. Mitochondrial H2O2 regulates the angiogenic phenotype via PTEN oxidation. J Biol Chem. 2005;280:16916–24. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
