

A novel mitochondrial sphingomyelinase in zebrafish cells
- PMID: 19429680
- PMCID: PMC2740460
- DOI: 10.1074/jbc.M109.004580
A novel mitochondrial sphingomyelinase in zebrafish cells
Abstract
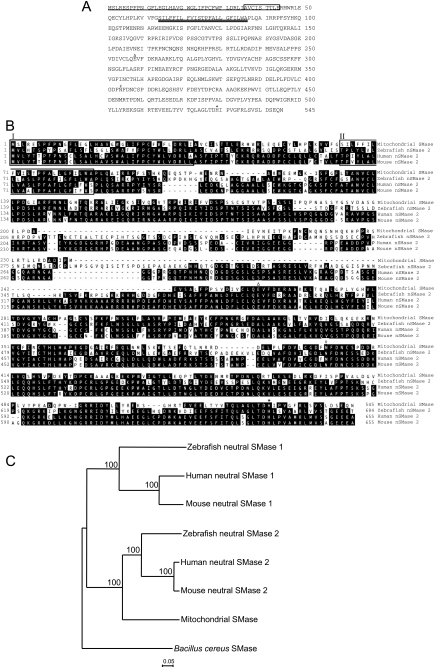
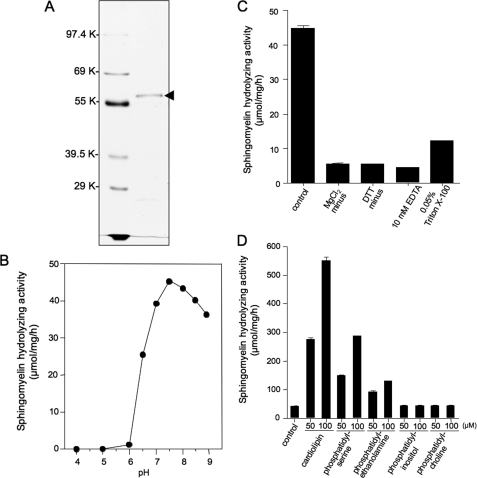
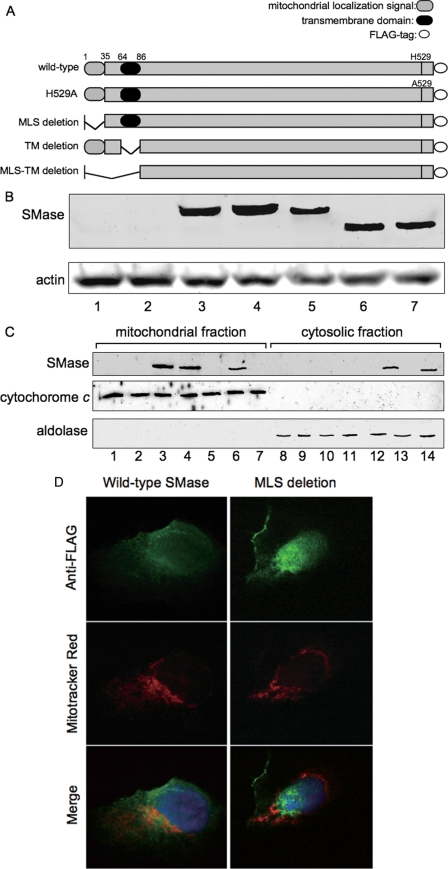
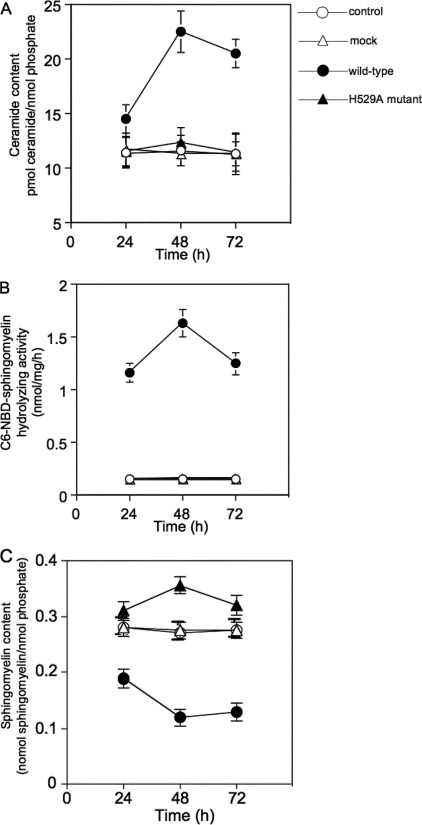
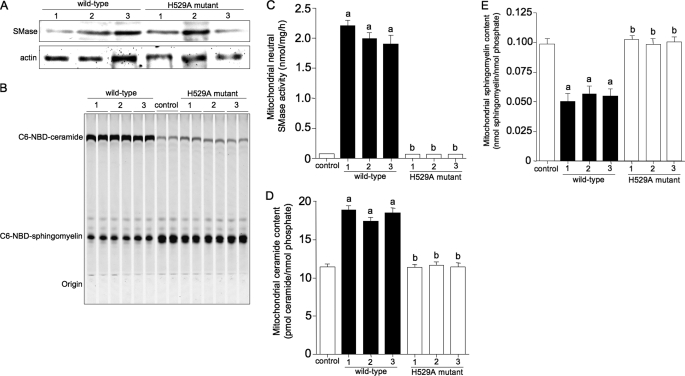
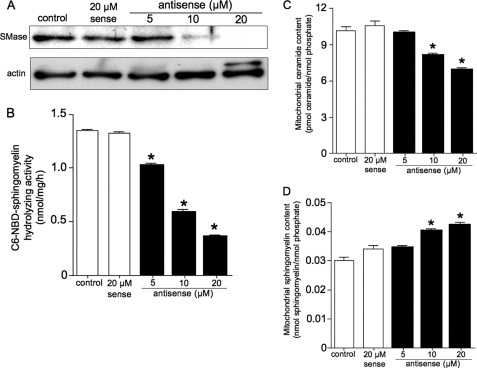
Sphingolipids are important signaling molecules in many biological processes, but little is known regarding their physiological roles in the mitochondrion. We focused on the biochemical characters of a novel sphingomyelinase (SMase) and its function in mitochondrial ceramide generation in zebrafish embryonic cells. The cloned SMase cDNA encoded a polypeptide of 545 amino acid residues (putative molecular weight, 61,300) containing a mitochondrial localization signal (MLS) and a predicted transmembrane domain. The mature endogenous enzyme was predicted to have a molecular weight of 57,000, and matrix-assisted laser de sorption ionization time-of-flight mass spectrometry analysis indicated that the N-terminal amino acid residue of the mature enzyme was Ala-36. The purified enzyme optimally hydrolyzed [(14)C]sphingomyelin in the presence of 10 mm Mg(2+) at pH 7.5. In HEK293 cells that overexpressed SMase cDNA, the enzyme was localized to the mitochondrial fraction, whereas mutant proteins lacking MLS or both the MLS and the transmembrane domain were absent from the mitochondrial fraction. Endogenous SMase protein co-localized with a mitochondrial cytostaining marker. Using a protease protection assay, we found that SMase was distributed throughout the intermembrane space and/or the inner membrane of the mitochondrion. Furthermore, the overexpression of SMase in HEK293 cells induced ceramide generation and sphingomyelin hydrolysis in the mitochondrial fraction. Antisense phosphorothioate oligonucleotide-induced knockdown repressed ceramide generation and sphingomyelin hydrolysis in the mitochondrial fraction in zebrafish embryonic cells. These observations indicate that SMase catalyzes the hydrolysis of sphingomyelin and generates ceramide in mitochondria in fish cells.
Figures

References
-
- Hannun Y. A., Luberto C. (2000) Trends Cell Biol. 10, 73–80 - PubMed
-
- Levade T., Malagarie-Cazenave S., Gouazé V., Ségui B., Tardy C., Betito S., Andrieu-Abadie N., Cuvillier O. (2002) Neurochem. Res. 27, 601–607 - PubMed
-
- Marchesini N., Hannun Y. A. (2004) Biochem. Cell Biol. 82, 27–44 - PubMed
-
- Obeid L. M., Linardic C. M., Karolak L. A., Hannun Y. A. (1993) Science 259, 1769–1771 - PubMed
-
- Okazaki T., Bell R. M., Hannun Y. A. (1989) J. Biol. Chem. 264, 19076–19080 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
