

Review
doi: 10.1242/dev.030403.
The function of non-coding RNAs in genomic imprinting
Affiliations
- PMID: 19429783
- PMCID: PMC2847617
- DOI: 10.1242/dev.030403
Item in Clipboard
Review
The function of non-coding RNAs in genomic imprinting
Development.
2009 Jun.
Abstract
Non-coding RNAs (ncRNAs) that regulate gene expression in cis or in trans are a shared feature of prokaryotic and eukaryotic genomes. In mammals, cis-acting functions are associated with macro ncRNAs, which can be several hundred thousand nucleotides long. Imprinted ncRNAs are well-studied macro ncRNAs that have cis-regulatory effects on multiple flanking genes. Recent advances indicate that they employ different downstream mechanisms to regulate gene expression in embryonic and placental tissues. A better understanding of these downstream mechanisms will help to improve our general understanding of the function of ncRNAs throughout the genome.
Figures
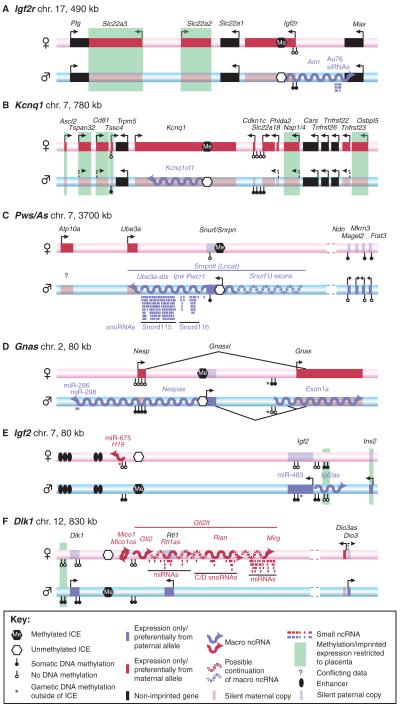
(A-F) The genomic organisation of six well-studied mouse imprinted clusters. The maternal chromosome is shown as a pink bar, the paternal chromosome as a blue bar. Protein-coding genes are shown as boxes: solid red box, maternally expressed gene on the expressed allele; transparent red box, maternally expressed gene on the repressed allele; solid blue box, paternally expressed gene on the expressed allele; transparent blue box, paternally expressed gene on the repressed allele. Macro ncRNAs are shown as wavy lines: red for maternally expressed, blue for paternally expressed. Arrows indicate transcriptional direction: solid arrows, strong transcription; dashed arrows, weak transcription. Note that many of the indicated genes show tissue- or temporal-restricted gene expression (not indicated). See key for further details. chr., chromosome; ICE, imprint control element; miRNA, microRNA; siRNA, short interfering RNA; snoRNA, short nucleolar RNA.
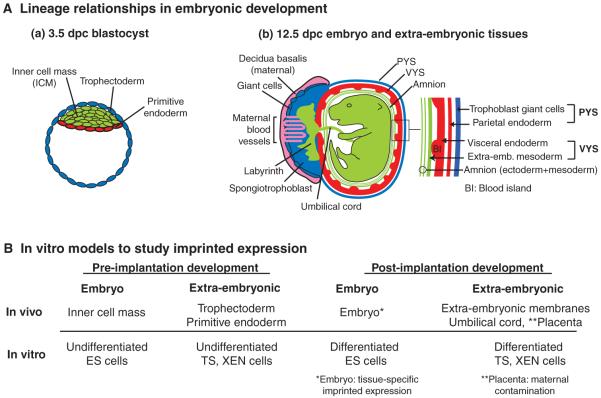
(A) Lineage relationships during embryonic development. (a) A blastocyst at 3.5 days post-coitum (dpc). The trophectoderm (blue) contributes to the parietal yolk sac (PYS) and the placenta of the 12.5 dpc embryo as depicted in b. The inner cell mass (ICM, green) gives rise to the embryo proper and contributes to the visceral yolk sac (VYS) and amnion extra-embryonic membranes. The primitive endoderm (red) differentiates into the endoderm layer of the PYS and VYS. (b) A 12.5 dpc embryo and its extra-embryonic tissues. The embryo (green) is surrounded by the amnion, which consists of ICM-derived ectoderm and mesoderm. The middle extra-embryonic membrane is the VYS, which consists of ICM-derived mesoderm (green) and endoderm (red). The outer membrane, the PYS, is lost after 13.5 dpc and consists of ICM-derived parietal endoderm (red) and trophoblast giant cells (blue), which are not ICM derived. The placenta consists of distinct layers: the inner labyrinth (green), the spongiotrophoblast (blue) and giant cells (blue). The outermost part of the placenta, the deciduas basalis (pink), is derived from maternal tissue. The intermingling of maternal blood vessels with the placenta is indicated. (B) In vitro model systems to study genomic imprinting. Undifferentiated embryonic stem (ES) cells are considered to mimic the blastocyst ICM, whereas undifferentiated trophoblast stem (TS) and extra-embryonic endoderm (XEN) cells mimic the blastocyst trophectoderm and primitive endoderm, respectively (Rossant, 2007). The differentiated derivatives of ES, TS and XEN cells might provide models for studying imprinting in post-implantation embryonic and extra-embryonic tissues, but care should be taken to ensure that the in vitro situations recapitulate what is observed in vivo.
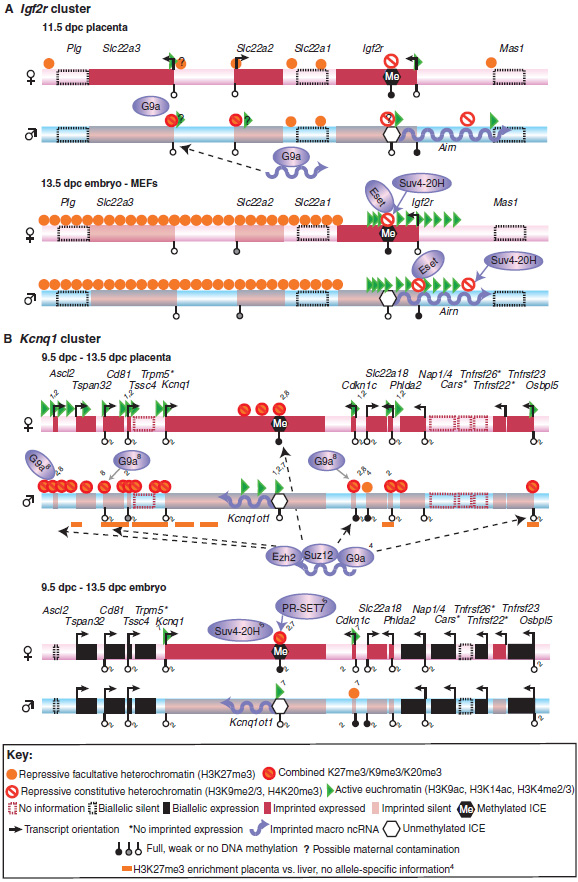
Active and repressive histone modifications on the maternal or paternal allele are shown for the (A) Igf2r and (B) Kcnq1 clusters for embryo and placenta. For simplicity, histone modifications are combined into three groups: repressive constitutive heterochromatin (H3K9me2/3, H4K20me3), repressive facultative heterochromatin (H3K27me3) and active euchromatin (H3K9ac, H3K4me2/3). In both clusters, only the indicated positions were assayed in the placenta, with the exception of one ChIP-chip mapping of H3K27me3 (orange lines). (A) In the Igf2r cluster, an unbiased continuous genome-wide location (ChIP-chip) analysis was performed in the embryo, with the indicated positions showing enrichment. (B) In the Kcnq1 cluster, all positions shown in the placenta were also assayed in the embryo, and only the indicated positions show allele-specific enrichment. Blue shaded ellipses indicate histone-modifying enzymes, and the solid blue arrows extending from the ellipses indicate the involvement of the respective enzyme in the setting or maintaining of a histone mark. Both Airn (A) and Kcnq1ot1 (B) ncRNAs bind to histone-modifying enzymes. Dashed arrows (B) indicate binding of Kcnq1ot1 ncRNA to chromatin. Gene expression marked by an asterisk is inferred from the presence of active histone modifications (Mikkelsen et al., 2007). References: Igf2r in the placenta (Nagano et al., 2008); Igf2r in the embryo (Regha et al., 2007); Kcnq1 in the placenta: all histone modifications without a numbered reference (Umlauf et al., 2004); others: 1(Green et al., 2007), 2(Lewis et al., 2004), 3(Mikkelsen et al., 2007), 4(Pandey et al., 2008), 5(Pannetier et al., 2008), 6(Regha et al., 2007), 7(Umlauf et al., 2004), 8(Wagschal et al., 2008). ac, acetylation; H3, histone 3; H4, histone 4; ICE, imprint control element; K, lysine; me2, dimethylation; me3, trimethylation.
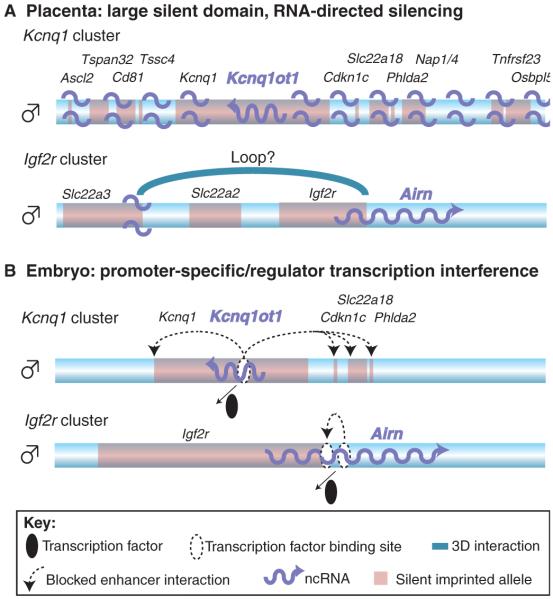
Imprinted macro ncRNAs might use different modes of silencing in embryonic and placental tissues. Only the paternal chromosome is shown (blue bar). The positions and names of silent imprinted genes are indicated in black font and by pale red boxes. Imprinted macro ncRNAs are indicated in blue font and by blue wavy arrows. (A) In the placenta, Kcnq1ot1 in the Kcnq1 cluster is transcribed from the paternal allele and localises to the whole imprinted domain by an unknown mechanism, inducing the recruitment of repressive histones, which leads to gene silencing. In the Igf2r cluster, Airn either locates to the silent Slc22a3 promoter, or the promoter forms a three-dimensional (3D) loop (indicated by a blue arch) to the Airn gene, similarly recruiting repressive histone modifications that silence Slc22a3. In both cases, the ncRNA itself is involved in the silencing process. (B) In embryonic tissues, only a few genes are silenced by the ncRNA in the Kcnq1 and Igf2r clusters. No localisation of the ncRNA to chromatin has been reported in the embryo. It is therefore possible that the transcription of the ncRNA is sufficient to silence all genes by interfering with the binding of essential transcription factors (black ellipse), thereby inducing gene silencing by interrupting enhancer interactions (dashed arrows).
References
-
- Allis CD, Berger SL, Cote J, Dent S, Jenuwien T, Kouzarides T, Pillus L, Reinberg D, Shi Y, Shiekhattar R, et al. New nomenclature for chromatin-modifying enzymes. Cell. 2007a;131:633–636. - PubMed
-
- Allis CD, Jenuwein T, Reinberg D. Overview and concepts. In: Allis CD, Jenuwein T, Reinberg D, Caparros M, editors. Epigenetics. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 2007b. pp. 23–61.
-
- Amaral PP, Mattick JS. Noncoding RNA in development. Mamm. Genome. 2008;19:454–492. - PubMed
-
- Barlow DP, Bartolomei MS. Genomic imprinting in mammals. In: Allis CD, Jenuwein T, Reinberg D, Caparros M, editors. Epigenetics. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 2007. pp. 357–375.
-
- Bell AC, Felsenfeld G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature. 2000;405:482–485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
