

The neural crest epithelial-mesenchymal transition in 4D: a 'tail' of multiple non-obligatory cellular mechanisms
- PMID: 19429784
- PMCID: PMC2680107
- DOI: 10.1242/dev.034785
The neural crest epithelial-mesenchymal transition in 4D: a 'tail' of multiple non-obligatory cellular mechanisms
Abstract
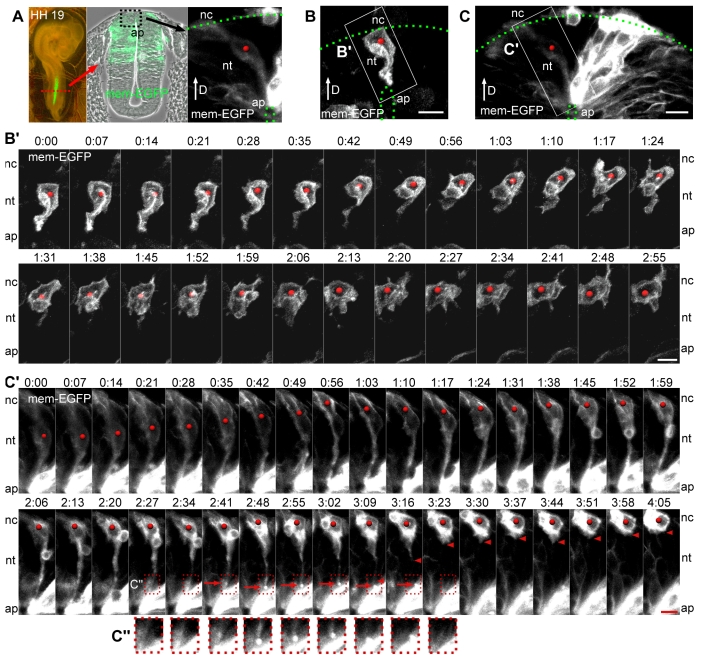
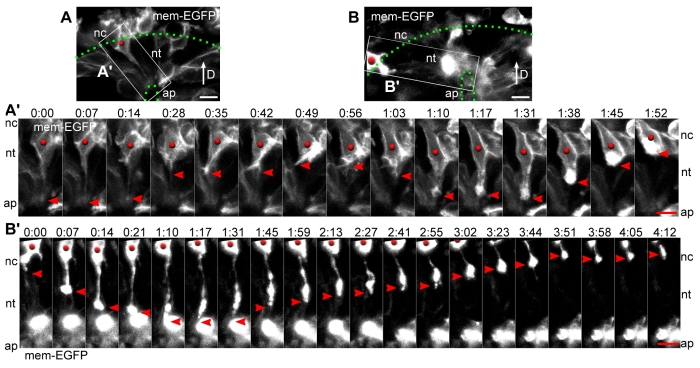
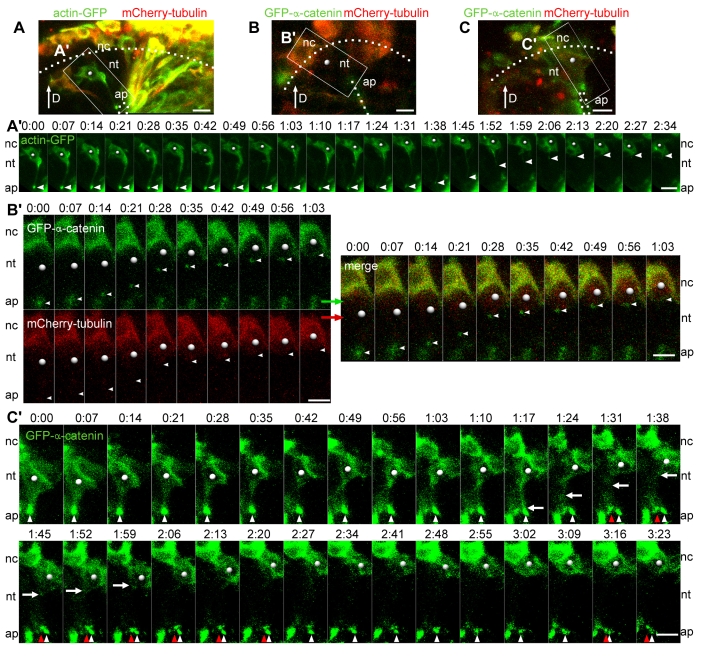
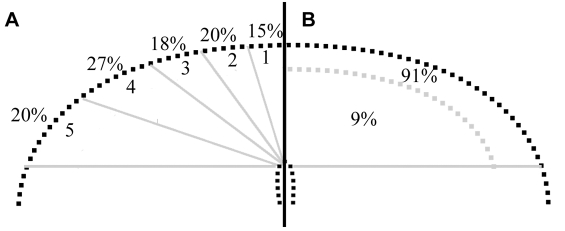
An epithelial-mesenchymal transition (EMT) is the process whereby epithelial cells become mesenchymal cells, and is typified by the generation of neural crest cells from the neuroepithelium of the dorsal neural tube. To investigate the neural crest EMT, we performed live cell confocal time-lapse imaging to determine the sequence of cellular events and the role of cell division in the EMT. It was observed that in most EMTs, the apical cell tail is retracted cleanly from the lumen of the neuroepithelium, followed by movement of the cell body out of the neural tube. However, exceptions to this sequence include the rupture of the neural crest cell tail during retraction (junctional complexes not completely downregulated), or translocation of the cell body away from the apical surface while morphologically rounded up in M phase (no cell tail retraction event). We also noted that cell tail retraction can occur either before or after the redistribution of apical-basolateral epithelial polarity markers. Surprisingly, we discovered that when an EMT was preceded by a mitotic event, the plane of cytokinesis does not predict neural crest cell fate. Moreover, when daughter cells are separated from the adherens junctions by a parallel mitotic cleavage furrow, most re-establish contact with the apical surface. The diversity of cellular mechanisms by which neural crest cells can separate from the neural tube suggests that the EMT program is a complex network of non-linear mechanisms that can occur in multiple orders and combinations to allow neural crest cells to escape from the neuroepithelium.
Figures



References
-
- Ahlstrom, J. D. and Erickson, C. A. (2007). Organization of cells into higher ordered structures. In Principles of Tissue Engineering (ed. R. Lanza, R. Langer and J. P. Vacanti), pp. 67-79. New York: Academic Press.
-
- Baum, B., Settleman, J. and Quinlan, M. P. (2008). Transitions between epithelial and mesenchymal states in development and disease. Semin. Cell Dev. Biol. 19, 294-308. - PubMed
-
- Bilozur, M. E. and Hay, E. D. (1989). Cell migration into neural tube lumen provides evidence for the “fixed cortex” theory of cell motility. Cell Motil. Cytoskeleton 14, 469-484. - PubMed
-
- Bronner-Fraser, M. and Fraser, S. E. (1988). Cell lineage analysis reveals multipotency of some avian neural crest cells. Nature 335, 161-164. - PubMed
-
- Burstyn-Cohen, T. and Kalcheim, C. (2002). Association between the cell cycle and neural crest delamination through specific regulation of G1/S transition. Dev. Cell 3, 383-395. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
