

Intraclonal competition limits the fate determination of regulatory T cells in the thymus
- PMID: 19430476
- PMCID: PMC2756247
- DOI: 10.1038/ni.1739
Intraclonal competition limits the fate determination of regulatory T cells in the thymus
Abstract
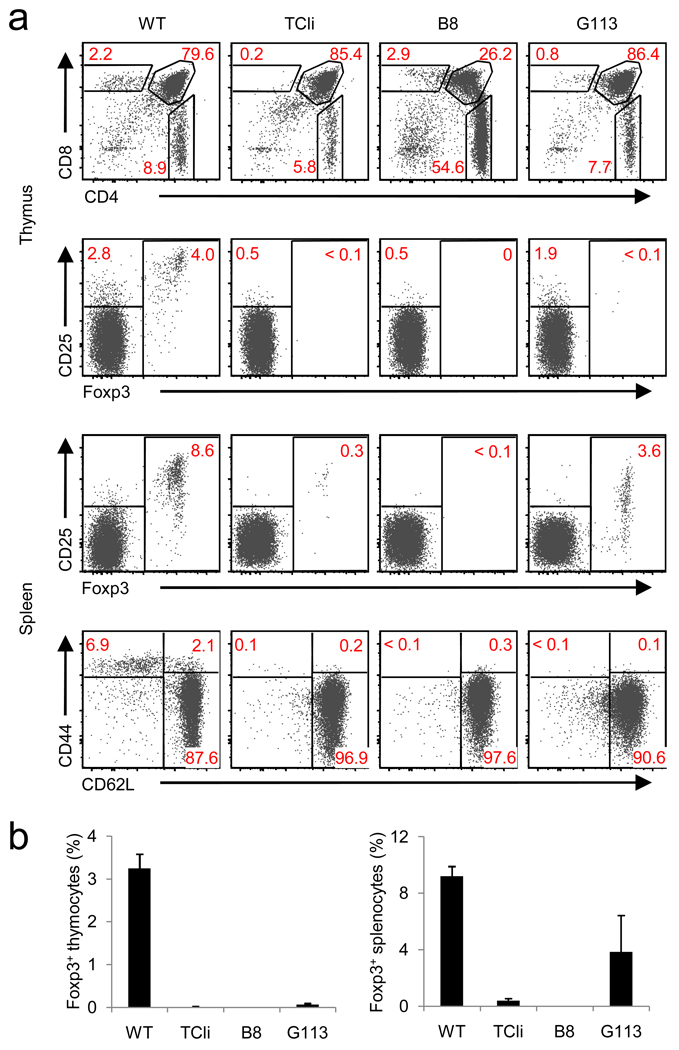
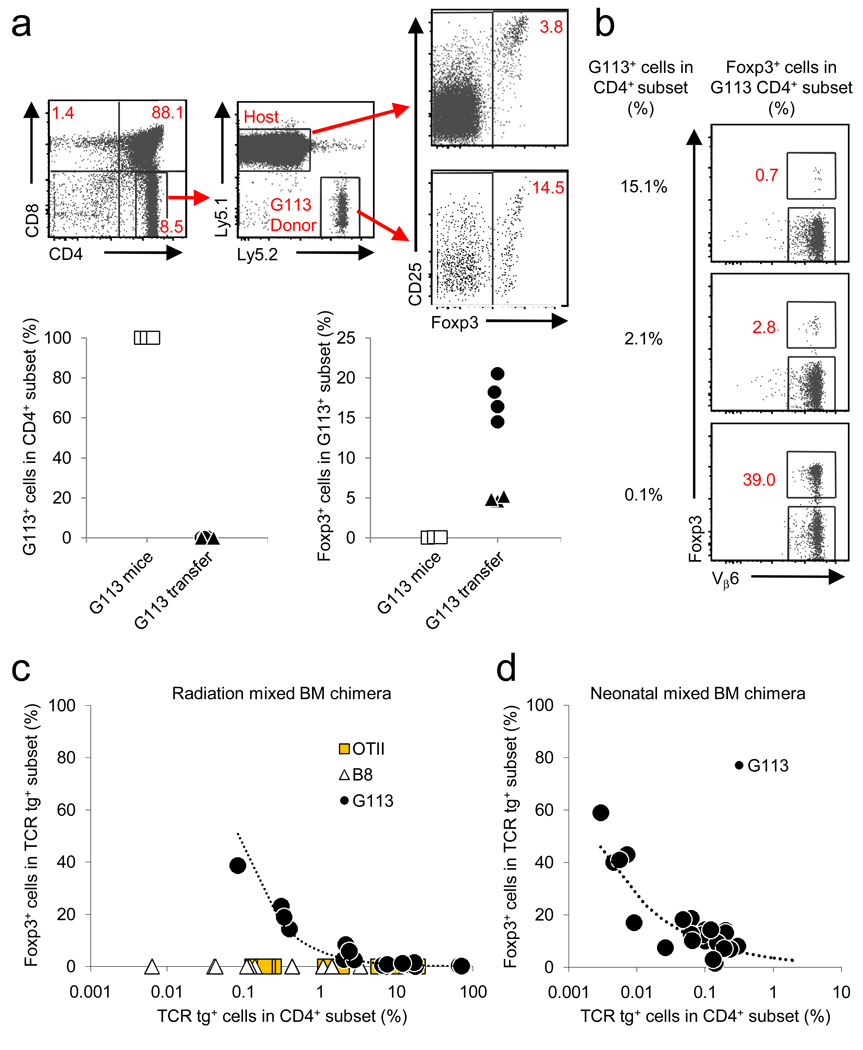
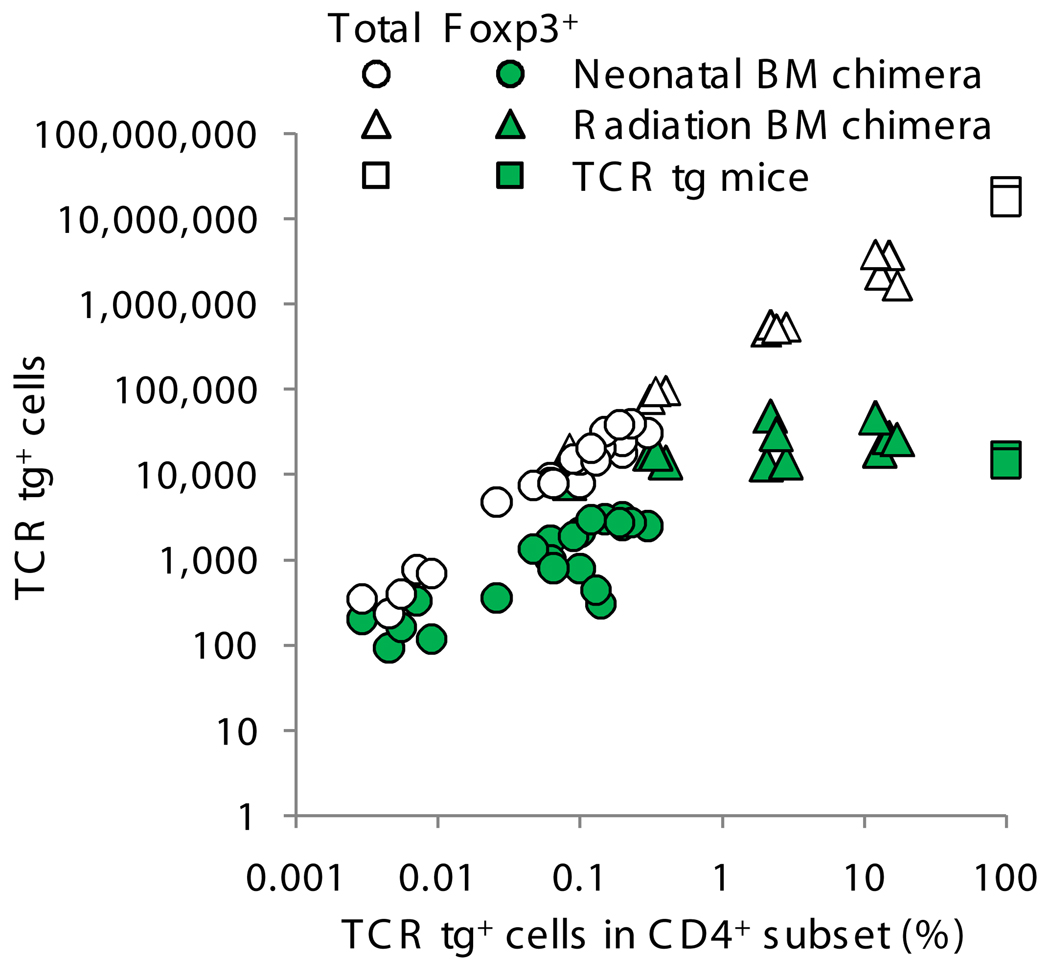
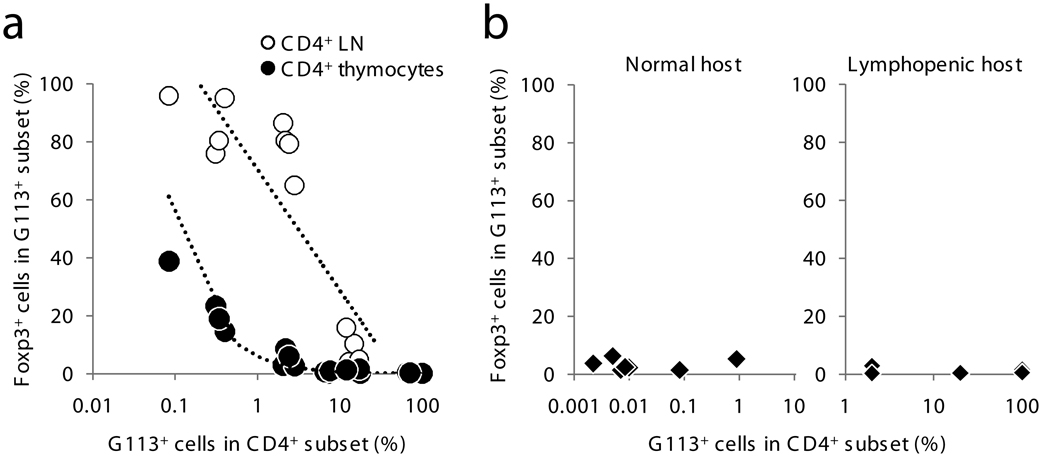
Because the deletion of self-reactive T cells is incomplete, thymic development of natural Foxp3+CD4+ regulatory T cells (Treg cells) is required for preventing autoimmunity. However, the function of T cell antigen receptor (TCR) specificity in thymic Treg cell development remains controversial. To address this issue, we generated a transgenic line expressing a naturally occurring Treg cell-derived TCR. Unexpectedly, we found that efficient thymic Treg cell development occurred only when the antigen-specific Treg cell precursors were present at low clonal frequency (o1%) in a normal thymus. Using retroviral vectors and bone marrow chimeras, we observed similar activity with two other Treg cell-derived TCRs. Our data demonstrate that thymic Treg cell development is a 'TCR-instructive' process involving a niche that can be saturable at much lower clonal frequencies than is the niche for positive selection.
Conflict of interest statement
The authors have no conflicts of interest to report.
Figures
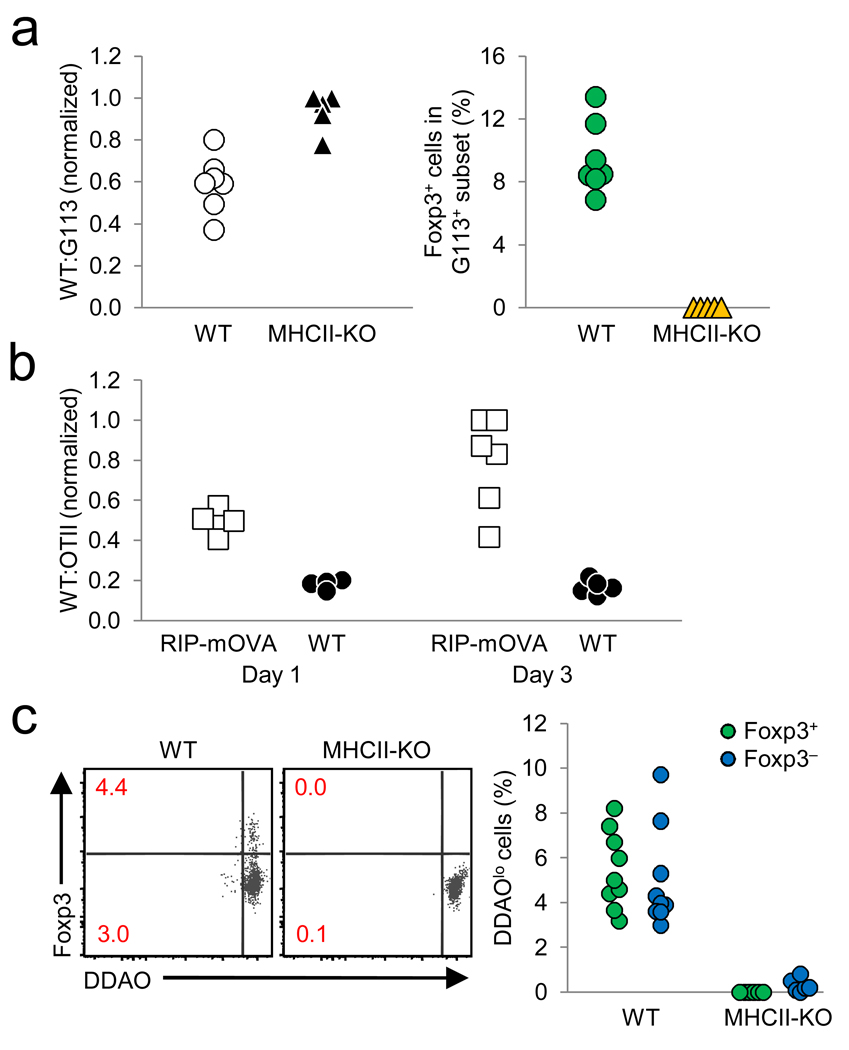
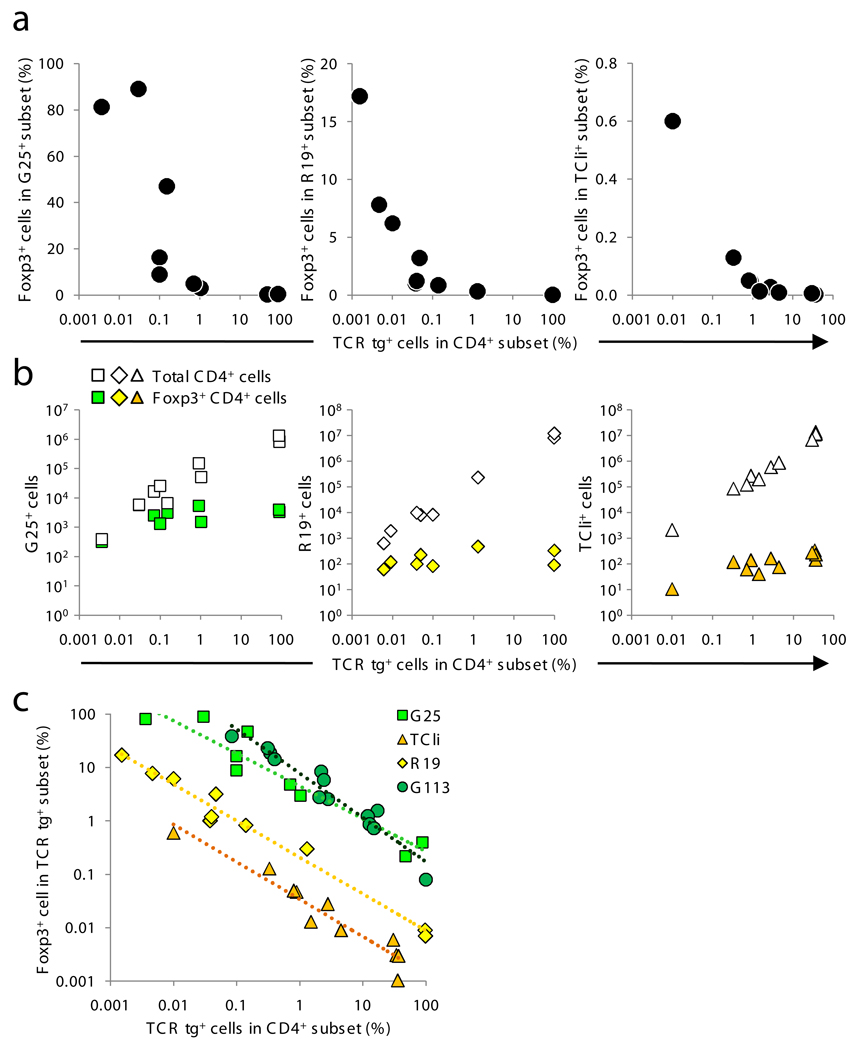
Comment in
-
Treg cells meet their limit.Nat Immunol. 2009 Jun;10(6):565-6. doi: 10.1038/ni0609-565. Nat Immunol. 2009. PMID: 19448655 No abstract available.
References
-
- Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133:775–787. - PubMed
-
- Kim JM, Rasmussen JP, Rudensky AY. Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nat. Immunol. 2007;8:191–197. - PubMed
-
- Curotto de Lafaille MA, et al. Adaptive Foxp3+ regulatory T cell-dependent and -independent control of allergic inflammation. Immunity. 2008;29:114–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
