

Decorin is a novel antagonistic ligand of the Met receptor
- PMID: 19433454
- PMCID: PMC2711571
- DOI: 10.1083/jcb.200901129
Decorin is a novel antagonistic ligand of the Met receptor
Abstract
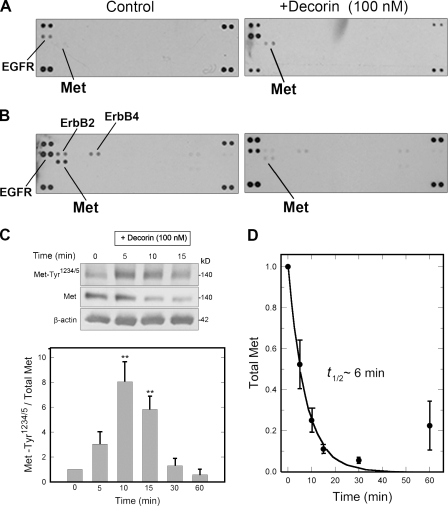
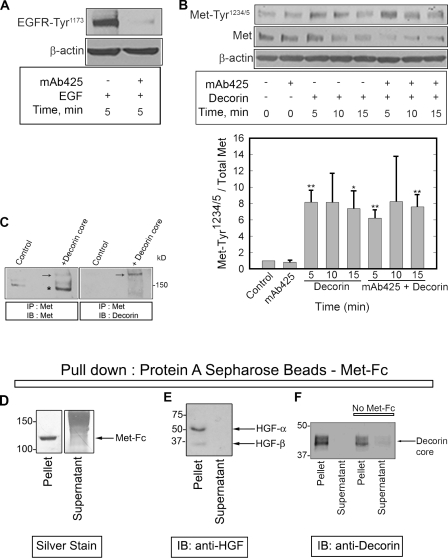
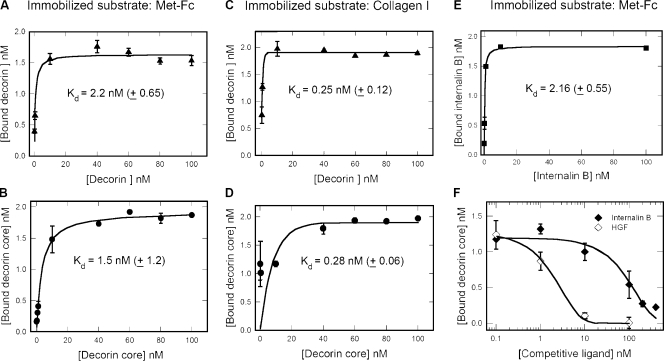
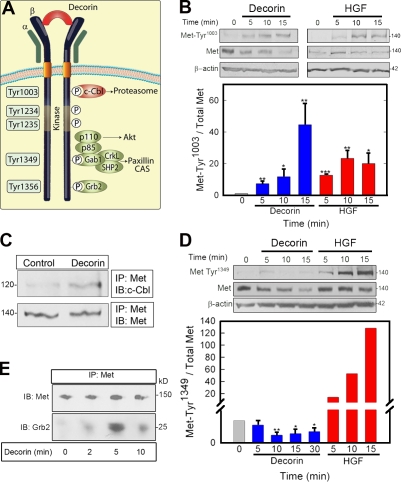
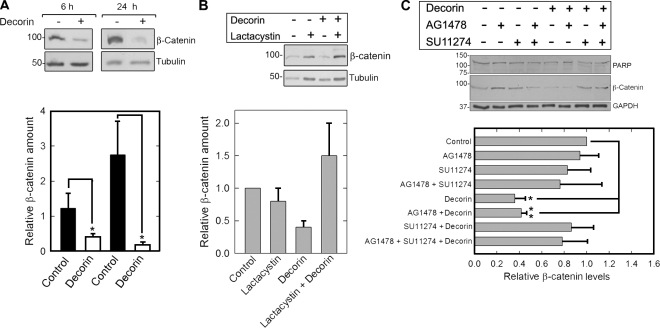
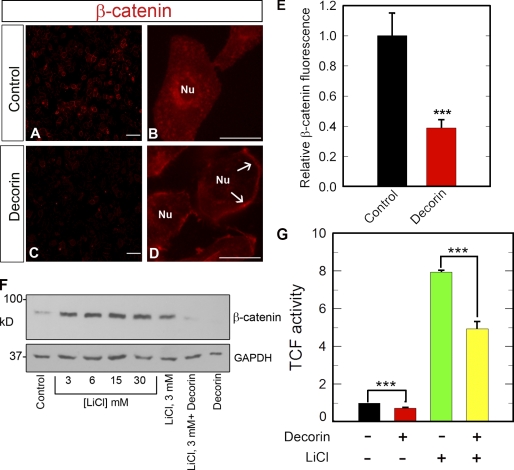
Decorin, a member of the small leucine-rich proteoglycan gene family, impedes tumor cell growth by down-regulating the epidermal growth factor receptor. Decorin has a complex binding repertoire, thus, we predicted that decorin would modulate the bioactivity of other tyrosine kinase receptors. We discovered that decorin binds directly and with high affinity (K(d) = approximately 1.5 nM) to Met, the receptor for hepatocyte growth factor (HGF). Binding of decorin to Met is efficiently displaced by HGF and less efficiently by internalin B, a bacterial Met ligand. Interaction of decorin with Met induces transient receptor activation, recruitment of the E3 ubiquitin ligase c-Cbl, and rapid intracellular degradation of Met (half-life = approximately 6 min). Decorin suppresses intracellular levels of beta-catenin, a known downstream Met effector, and inhibits Met-mediated cell migration and growth. Thus, by antagonistically targeting multiple tyrosine kinase receptors, decorin contributes to reduction in primary tumor growth and metastastic spreading.
Figures
References
-
- Athauda G., Giubellino A., Coleman J.A., Horak C., Steeg P.S., Lee M.-J., Trepel J., Wimberly J., Sun J., Coxon A., et al. 2006. c-Met ectodomain shedding rate correlates with malignant potential.Clin. Cancer Res. 12:4154–4162 - PubMed
-
- Berthou S., Aebersold D.M., Schimdt L.S., Stroka D., Heigl C., Streit B., Stalder D., Gruber G., Liang S., Howlett A.R., et al. 2004. The Met kinase inhibitor SU11274 exhibits a selective inhibition pattern toward different receptor mutated variants.Oncogene. 23:5387–5393 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
