

Gating, permselectivity and pH-dependent modulation of channels formed by connexin57, a major connexin of horizontal cells in the mouse retina
- PMID: 19433576
- PMCID: PMC2727035
- DOI: 10.1113/jphysiol.2009.171496
Gating, permselectivity and pH-dependent modulation of channels formed by connexin57, a major connexin of horizontal cells in the mouse retina
Abstract
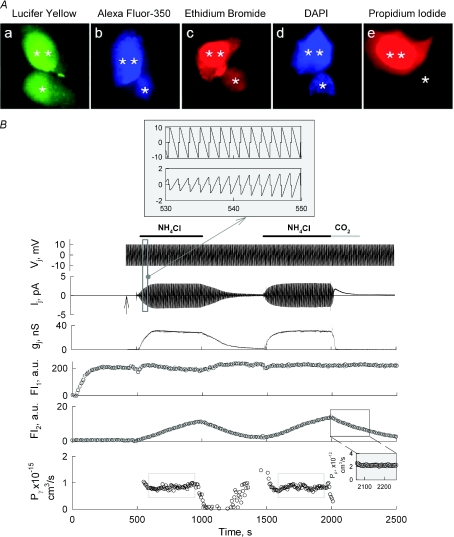
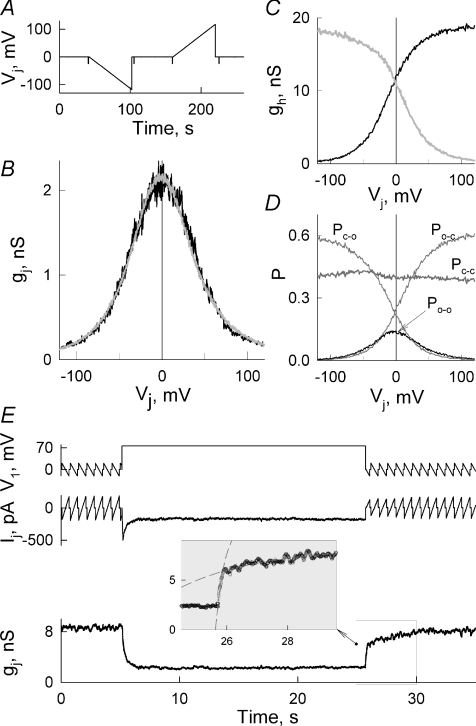
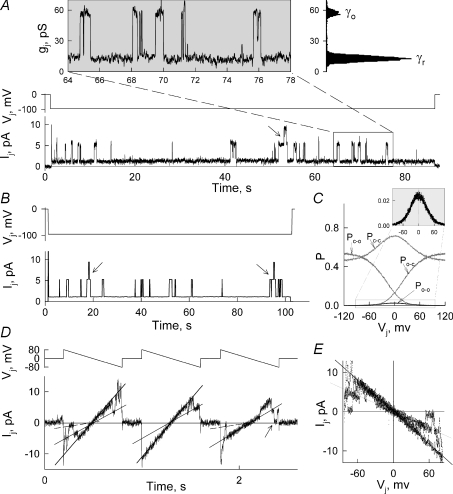
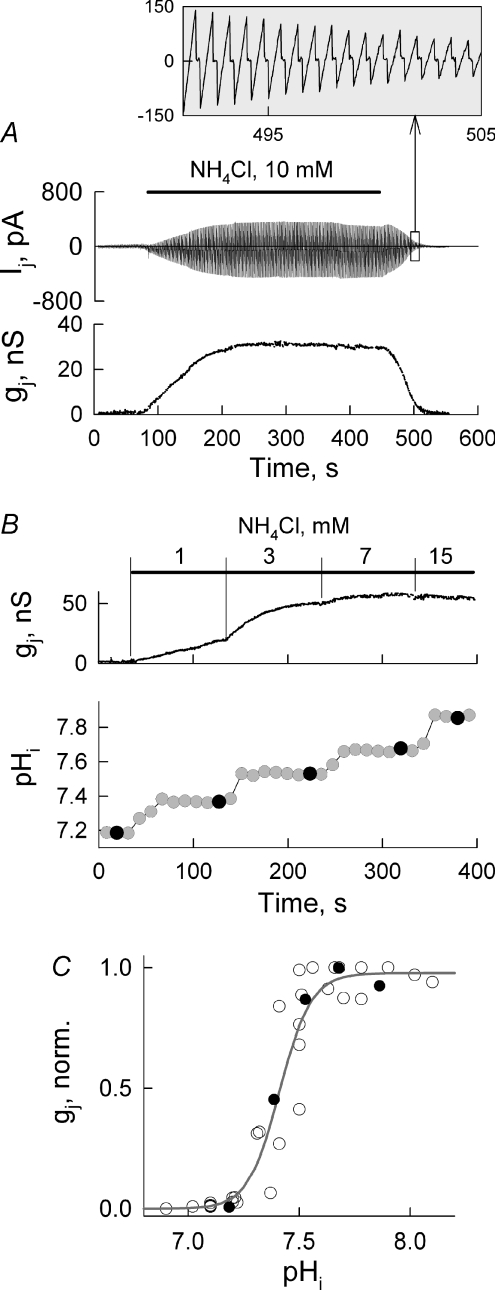
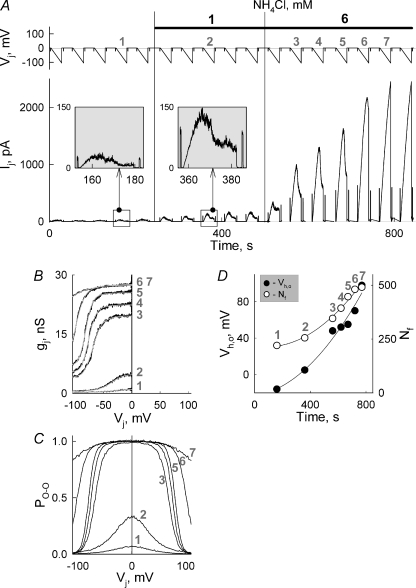
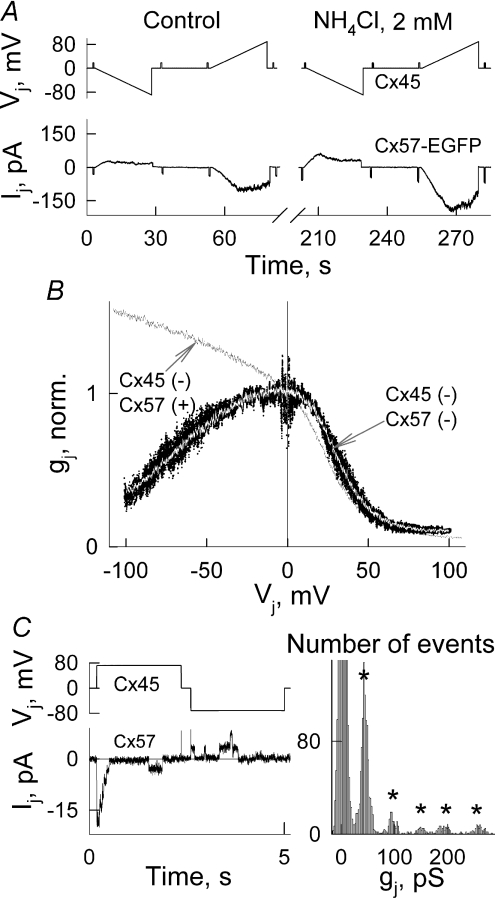
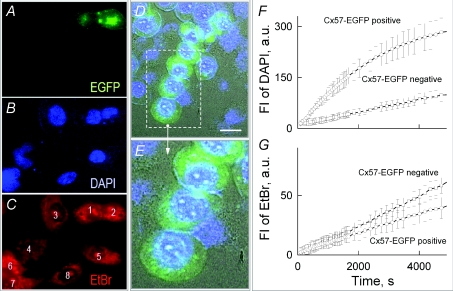
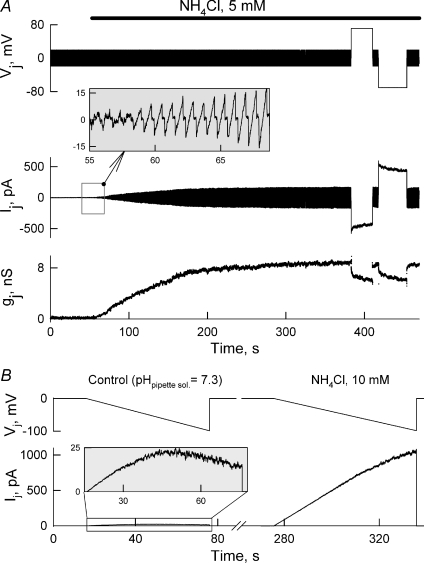
Mouse connexin57 (Cx57) is expressed most abundantly in horizontal cells of the retina, and forms gap junction (GJ) channels, which constitute a structural basis for electrical and metabolic intercellular communication, and unapposed hemichannels (UHCs) that are involved in an exchange of ions and metabolites between the cytoplasm and extracellular milieu. By combining fluorescence imaging and dual whole-cell voltage clamp methods, we showed that HeLa cells expressing Cx57 and C-terminally fused with enhanced green fluorescent protein (Cx57-EGFP) form junctional plaques (JPs) and that only cell pairs exhibiting at least one JP demonstrate cell-to-cell electrical coupling and transfer of negatively and positively charged dyes with molecular mass up to approximately 400 Da. The permeability of the single Cx57 GJ channel to Alexa fluor-350 is approximately 90-fold smaller than the permeability of Cx43, while its single channel conductance (57 pS) is only 2-fold smaller than Cx43 (110 pS). Gating of Cx57-EGFP/Cx45 heterotypic GJ channels reveal that Cx57 exhibit a negative gating polarity, i.e. channels tend to close at negativity on the cytoplasmic side of Cx57. Alkalization of pH(i) from 7.2 to 7.8 increased gap junctional conductance (g(j)) of approximately 100-fold with pK(a) = 7.41. We show that this g(j) increase was caused by an increase of both the open channel probability and the number of functional channels. Function of Cx57 UHCs was evaluated based on the uptake of fluorescent dyes. We found that under control conditions, Cx57 UHCs are closed and open at [Ca(2+)](o) = approximately 0.3 mm or below, demonstrating that a moderate reduction of [Ca(2+)](o) can facilitate the opening of Cx57 UHCs. This was potentiated with intracellular alkalization. In summary, our data show that the open channel probability of Cx57 GJs can be modulated by pH(i) with very high efficiency in the physiologically relevant range and may explain pH-dependent regulation of cell-cell coupling in horizontal cell in the retina.
Figures
References
-
- Baldridge WH, Ball AK, Miller RG. Dopaminergic regulation of horizontal cell gap junction particle density in goldfish retina. J Comp Neurol. 1987;265:428–436. - PubMed
-
- Becker D, Bonness VV, Mobbs P. Cell coupling in the retina: patterns and purpose. Cell Biol Intern. 1998;22:781–792. - PubMed
-
- Bennett MV, Barrio LC, Bargiello TA, Spray DC, Hertzberg E, Saez JC. Gap junctions: new tools, new answers, new questions. Neuron. 1991;6:305–320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous