

A hotspot of inactivation: The A22S and V108M polymorphisms individually destabilize the active site structure of catechol O-methyltransferase
- PMID: 19435324
- PMCID: PMC2906713
- DOI: 10.1021/bi900174v
A hotspot of inactivation: The A22S and V108M polymorphisms individually destabilize the active site structure of catechol O-methyltransferase
Abstract
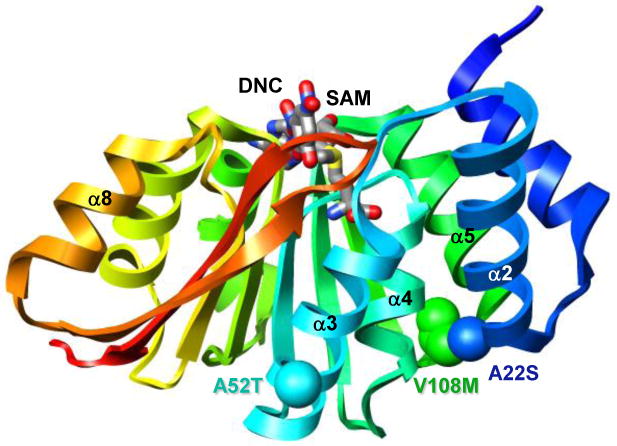
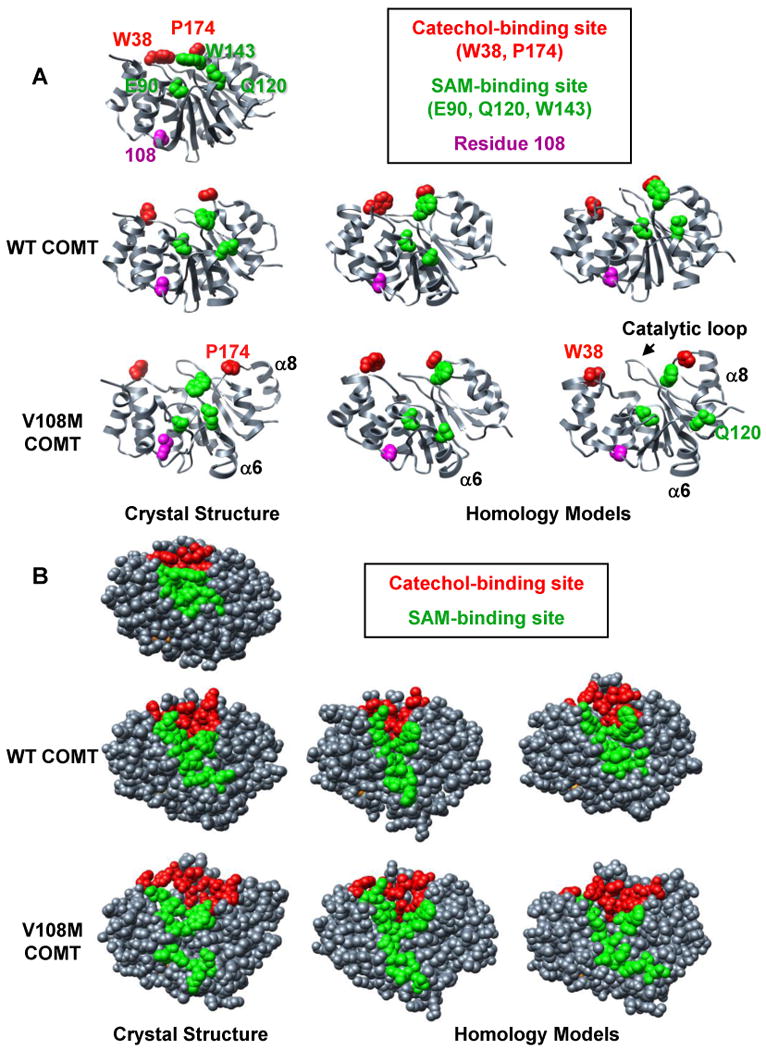
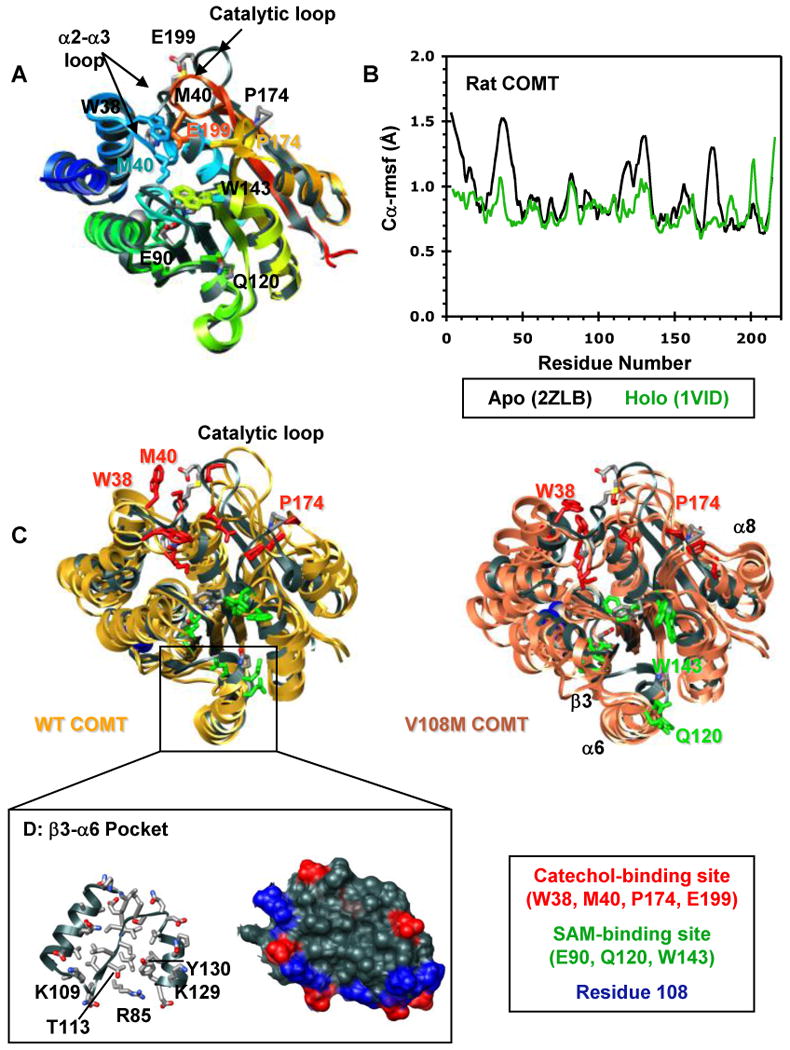
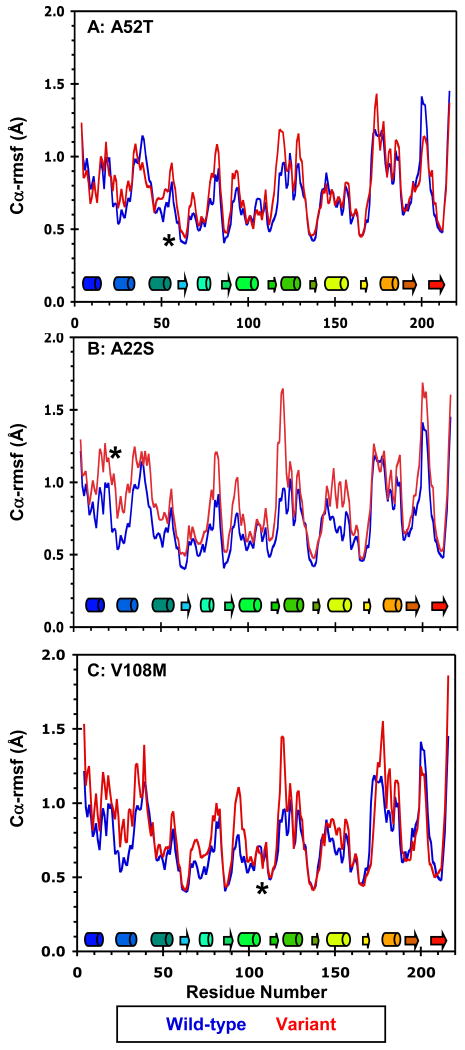
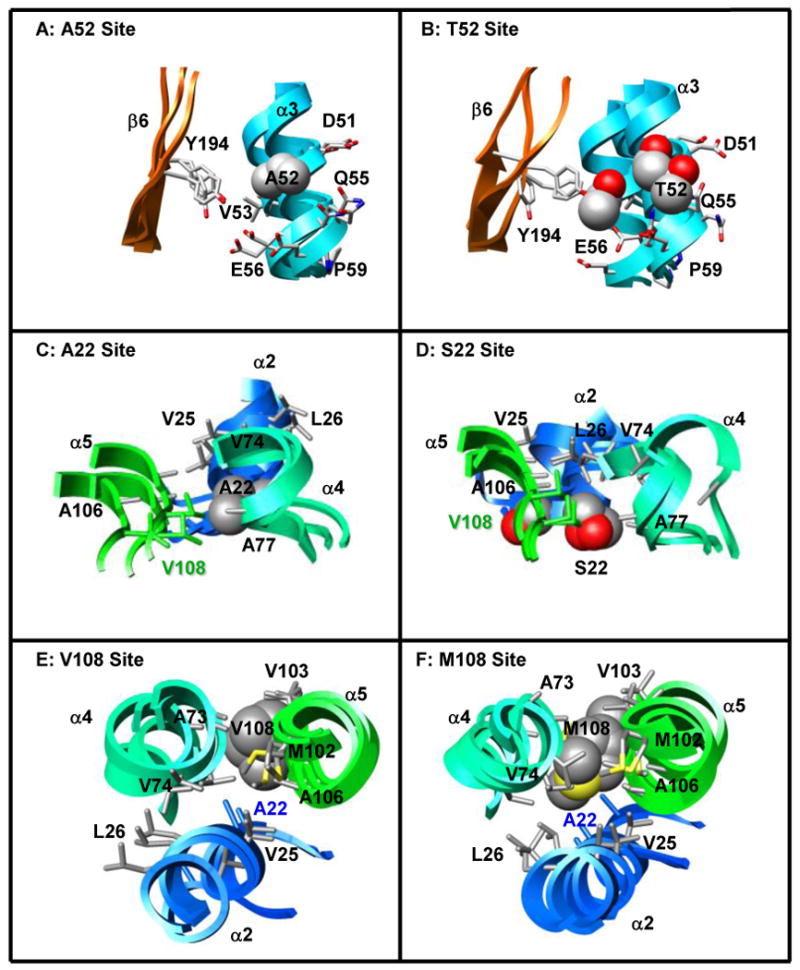
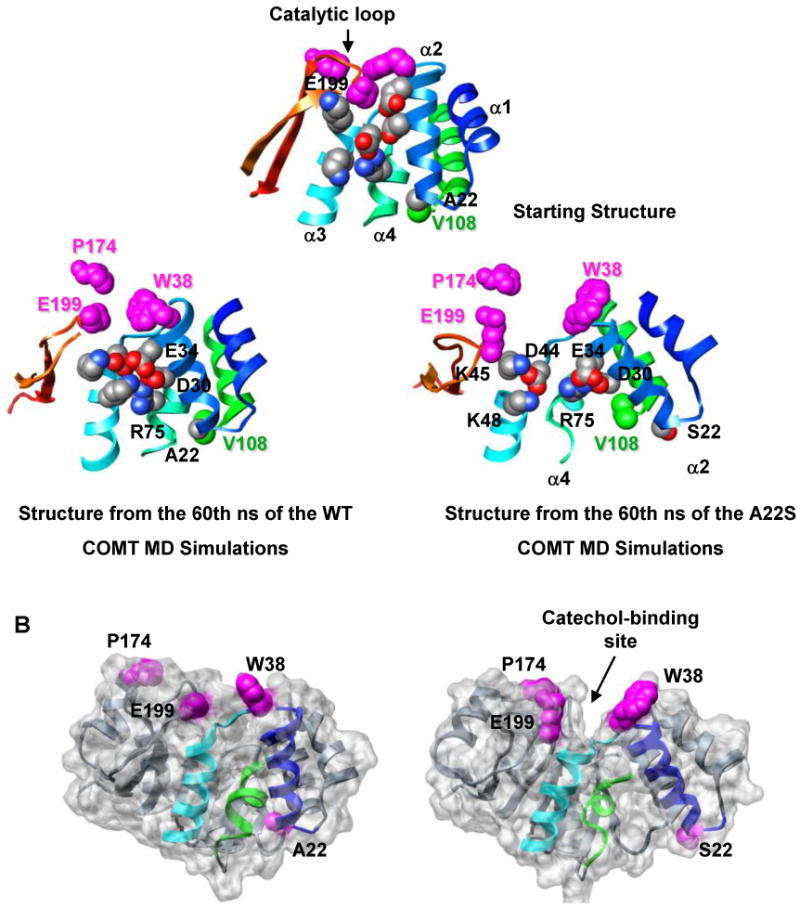
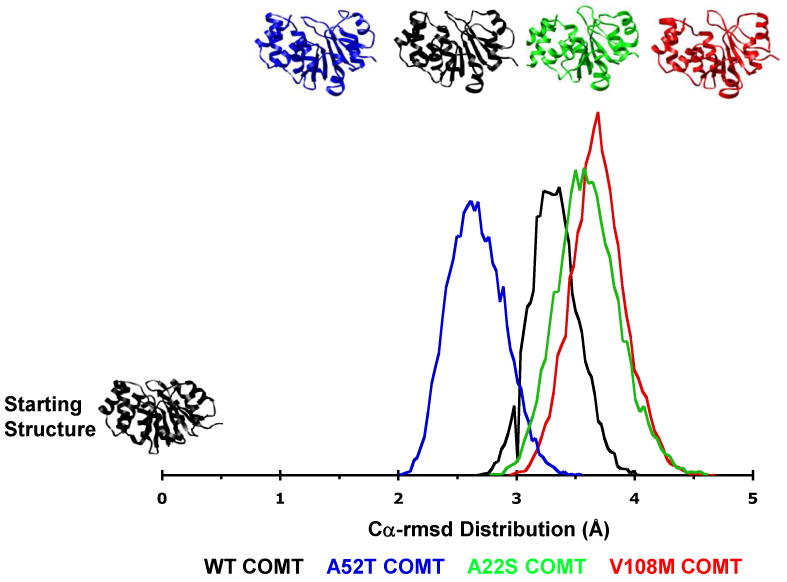
Human catechol O-methyltransferase (COMT) contains three common polymorphisms (A22S, A52T, and V108M), two of which (A22S and V108M) render the protein susceptible to deactivation by temperature or oxidation. We have performed multiple molecular dynamics simulations of the wild-type, A22S, A52T, and V108M COMT proteins to explore the structural consequences of these mutations. In total, we have amassed more than 1.4 micros of simulation time, representing the largest set of simulations detailing the effects of polymorphisms on a protein system to date. The A52T mutation had no significant effect on COMT structure in accord with experiment, thereby serving as a good negative control for the simulation set. Residues 22 (alpha2) and 108 (alpha5) interact with each other throughout the simulations and are located in a polymorphic hotspot approximately 20 A from the active site. Introduction of either the larger Ser (22) or Met (108) tightens this interaction, pulling alpha2 and alpha5 toward each other and away from the protein core. The V108M polymorphism rearranges active-site residues in alpha5, beta3, and alpha6, increasing the S-adenosylmethionine site solvent exposure. The A22S mutation reorients alpha2, moving critical catechol-binding residues away from the substrate-binding pocket. The A22S and V108M polymorphisms evolved independently in Northern European and Asian populations. While the decreased activities of both A22S and V108M COMT are associated with an increased risk for schizophrenia, the V108M-induced destabilization is also linked with improved cognitive function. These results suggest that polymorphisms within this hotspot may have evolved to regulate COMT activity and that heterozygosity for either mutation may be advantageous.
Figures
References
-
- Axelrod J, Szara S. Enzymic conversion of metanephrine to epinephrine. Biochim Biophys Acta. 1958;30:188–189. - PubMed
-
- Mannisto PT, Kaakkola S. Catechol-O-methyltransferase (COMT): biochemistry, molecular biology, pharmacology, and clinical efficacy of the new selective COMT inhibitors. Pharmacol Rev. 1999;51:593–628. - PubMed
-
- Weinshilboum RM, Otterness DM, Szumlanski CL. Methylation pharmacogenetics: catechol O-methyltransferase, thiopurine methyltransferase, and histamine N-methyltransferase. Annu Rev Pharmacol Toxicol. 1999;39:19–52. - PubMed
-
- Tenhunen J, Salminen M, Jalanko A, Ukkonen S, Ulmanen I. Structure of the rat catechol-O-methyltransferase gene: separate promoters are used to produce mRNAs for soluble and membrane-bound forms of the enzyme. DNA Cell Biol. 1993;12:253–263. - PubMed
-
- Tenhunen J, Salminen M, Lundstrom K, Kiviluoto T, Savolainen R, Ulmanen I. Genomic organization of the human catechol O-methyltransferase gene and its expression from two distinct promoters. Eur J Biochem. 1994;223:1049–1059. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
