

PfeIK1, a eukaryotic initiation factor 2alpha kinase of the human malaria parasite Plasmodium falciparum, regulates stress-response to amino-acid starvation
- PMID: 19435497
- PMCID: PMC2684119
- DOI: 10.1186/1475-2875-8-99
PfeIK1, a eukaryotic initiation factor 2alpha kinase of the human malaria parasite Plasmodium falciparum, regulates stress-response to amino-acid starvation
Abstract
Background: Post-transcriptional control of gene expression is suspected to play an important role in malaria parasites. In yeast and metazoans, part of the stress response is mediated through phosphorylation of eukaryotic translation initiation factor 2alpha (eIF2alpha), which results in the selective translation of mRNAs encoding stress-response proteins.
Methods: The impact of starvation on the phosphorylation state of PfeIF2alpha was examined. Bioinformatic methods were used to identify plasmodial eIF2alpha kinases. The activity of one of these, PfeIK1, was investigated using recombinant protein with non-physiological substrates and recombinant PfeIF2alpha. Reverse genetic techniques were used to disrupt the pfeik1 gene.
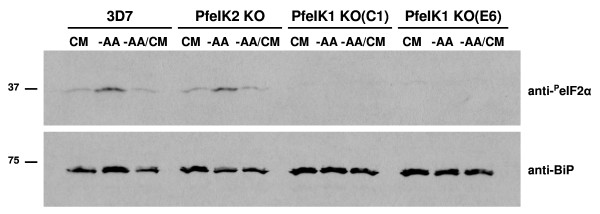
Results: The data demonstrate that the Plasmodium falciparum eIF2alpha orthologue is phosphorylated in response to starvation, and provide bioinformatic evidence for the presence of three eIF2alpha kinases in P. falciparum, only one of which (PfPK4) had been described previously. Evidence is provided that one of the novel eIF2alpha kinases, PfeIK1, is able to phosphorylate the P. falciparum eIF2alpha orthologue in vitro. PfeIK1 is not required for asexual or sexual development of the parasite, as shown by the ability of pfeik1- parasites to develop into sporozoites. However, eIF2alpha phosphorylation in response to starvation is abolished in pfeik1- asexual parasites
Conclusion: This study strongly suggests that a mechanism for versatile regulation of translation by several kinases with a similar catalytic domain but distinct regulatory domains, is conserved in P. falciparum.
Figures






Similar articles
-
Molecular cloning, characterization and localization of PfPK4, an eIF-2alpha kinase-related enzyme from the malarial parasite Plasmodium falciparum.Biochem J. 1997 Dec 1;328 ( Pt 2)(Pt 2):677-87. doi: 10.1042/bj3280677. Biochem J. 1997. PMID: 9371731 Free PMC article.
-
The Adaptive Proline Response in P. falciparum Is Independent of PfeIK1 and eIF2α Signaling.ACS Infect Dis. 2019 Apr 12;5(4):515-520. doi: 10.1021/acsinfecdis.8b00363. Epub 2019 Feb 21. ACS Infect Dis. 2019. PMID: 30773881 Free PMC article.
-
PKR and GCN2 kinases and guanine nucleotide exchange factor eukaryotic translation initiation factor 2B (eIF2B) recognize overlapping surfaces on eIF2alpha.Mol Cell Biol. 2005 Apr;25(8):3063-75. doi: 10.1128/MCB.25.8.3063-3075.2005. Mol Cell Biol. 2005. PMID: 15798194 Free PMC article.
-
Translational control in Plasmodium and toxoplasma parasites.Eukaryot Cell. 2013 Feb;12(2):161-7. doi: 10.1128/EC.00296-12. Epub 2012 Dec 14. Eukaryot Cell. 2013. PMID: 23243065 Free PMC article. Review.
-
The integrated stress response in metabolic adaptation.J Biol Chem. 2024 Apr;300(4):107151. doi: 10.1016/j.jbc.2024.107151. Epub 2024 Mar 9. J Biol Chem. 2024. PMID: 38462161 Free PMC article. Review.
Cited by
-
The role of upstream open reading frames in translation regulation in the apicomplexan parasites Plasmodium falciparum and Toxoplasma gondii.Parasitology. 2021 Sep;148(11):1277-1287. doi: 10.1017/S0031182021000937. Epub 2021 Jun 8. Parasitology. 2021. PMID: 34099078 Free PMC article. Review.
-
The cytoplasmic prolyl-tRNA synthetase of the malaria parasite is a dual-stage target of febrifugine and its analogs.Sci Transl Med. 2015 May 20;7(288):288ra77. doi: 10.1126/scitranslmed.aaa3575. Sci Transl Med. 2015. PMID: 25995223 Free PMC article.
-
A non-canonical sensing pathway mediates Plasmodium adaptation to amino acid deficiency.Commun Biol. 2023 Feb 21;6(1):205. doi: 10.1038/s42003-023-04566-y. Commun Biol. 2023. PMID: 36810637 Free PMC article.
-
A GCN2-Like eIF2α Kinase (LdeK1) of Leishmania donovani and Its Possible Role in Stress Response.PLoS One. 2016 Jun 1;11(6):e0156032. doi: 10.1371/journal.pone.0156032. eCollection 2016. PLoS One. 2016. PMID: 27248816 Free PMC article.
-
Autophagy in Plasmodium, a multifunctional pathway?Comput Struct Biotechnol J. 2013 Aug 20;8:e201308002. doi: 10.5936/csbj.201308002. eCollection 2013. Comput Struct Biotechnol J. 2013. PMID: 24688742 Free PMC article. Review.
References
-
- Hall N, Karras M, Raine JD, Carlton JM, Kooij TW, Berriman M, Florens L, Janssen CS, Pain A, Christophides GK, James K, Rutherford K, Harris B, Harris D, Churcher C, Quail MA, Ormond D, Doggett J, Trueman HE, Mendoza J, Bidwell SL, Rajandream MA, Carucci DJ, Yates JR, 3rd, Kafatos FC, Janse CJ, Barrell B, Turner CM, Waters AP, Sinden RE. A comprehensive survey of the Plasmodium life cycle by genomic, transcriptomic, and proteomic analyses. Science. 2005;307:82–86. doi: 10.1126/science.1103717. - DOI - PubMed
-
- Paton MG, Barker GC, Matsuoka H, Ramesar J, Janse CJ, Waters AP, Sinden RE. Structure and expression of a post-transcriptionally regulated malaria gene encoding a surface protein from the sexual stages of Plasmodium berghei. Mol Biochem Parasitol. 1993;59:263–275. doi: 10.1016/0166-6851(93)90224-L. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
