

Chibby forms a homodimer through a heptad repeat of leucine residues in its C-terminal coiled-coil motif
- PMID: 19435523
- PMCID: PMC2686680
- DOI: 10.1186/1471-2199-10-41
Chibby forms a homodimer through a heptad repeat of leucine residues in its C-terminal coiled-coil motif
Abstract
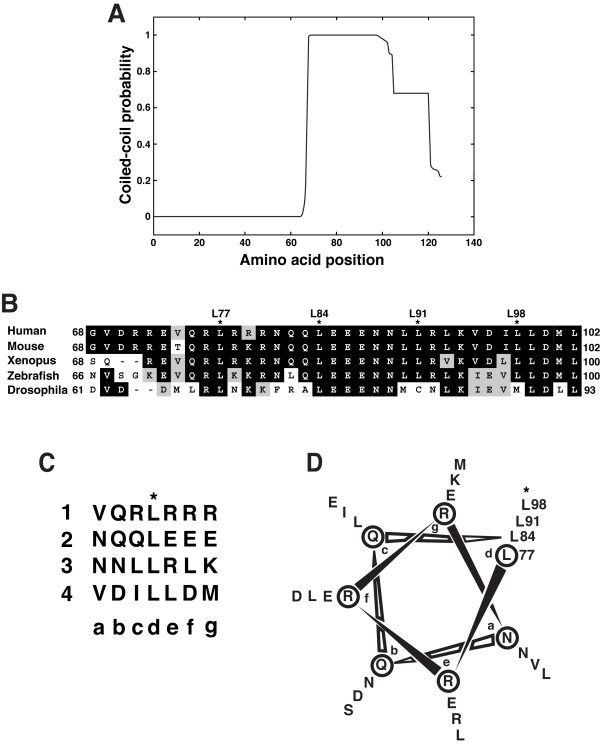
Background: The Wnt/beta-catenin signaling pathway plays crucial roles in embryonic development and in maintenance of organs and tissues in adults. Chibby (Cby) is an evolutionarily conserved molecule that physically interacts with the key downstream coactivator beta-catenin and represses its transcriptional activation potential. Although Cby harbors a predicted coiled-coil motif in the C-terminal region, its molecular nature and functional importance remain largely unexplored.
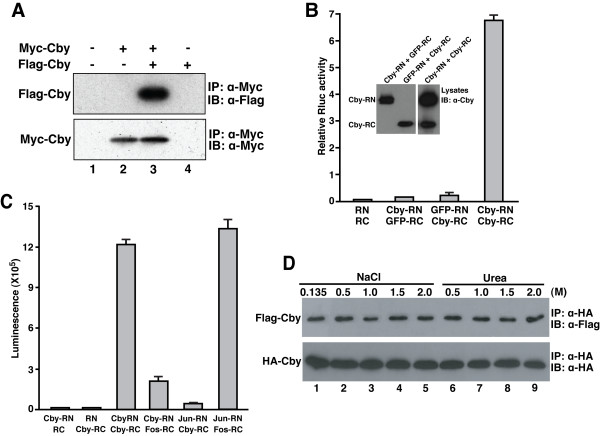
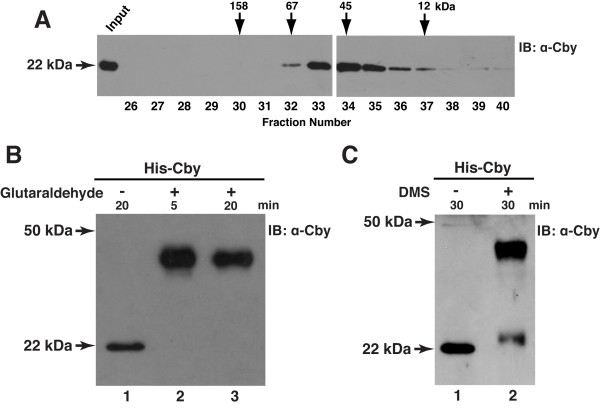
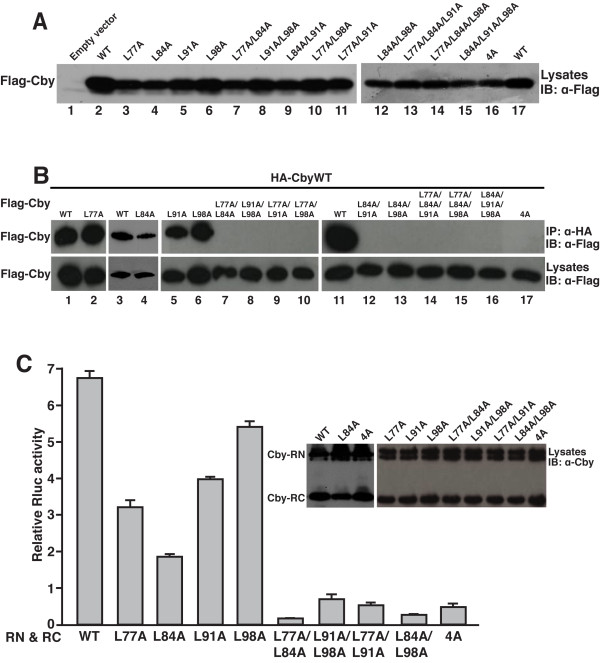
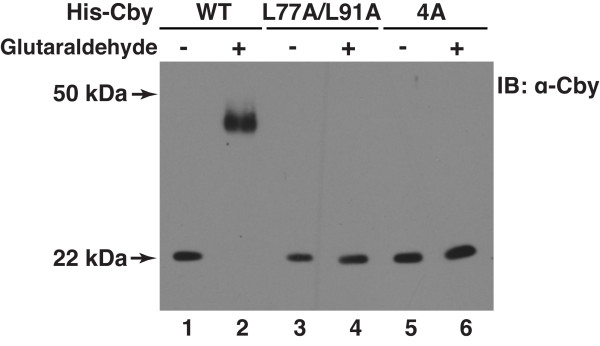
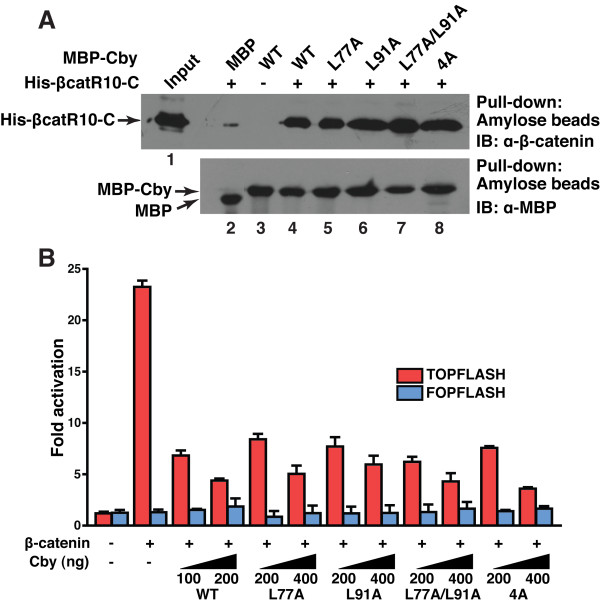
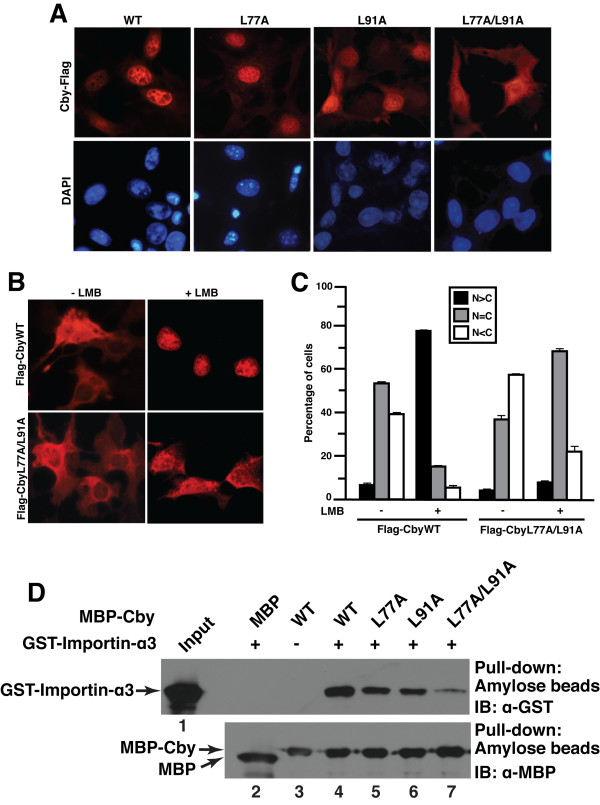
Results: Here we report that Cby forms a stable complex with itself. Alanine substitutions of two or more of four critical leucine residues within the C-terminal heptad repeats completely eliminate the Cby-Cby interaction. The Cby oligomer predominantly exists as a homodimer. Furthermore, we found that dimerization-deficient Cby mutants still retain the ability to bind to beta-catenin and to repress beta-catenin-dependent gene activation. More importantly, Cby homodimerization is required for its efficient interaction with the nuclear import receptor importin-alpha and subsequent nuclear translocation.
Conclusion: Our comprehensive mutational analysis of the Cby coiled-coil domain reveals that the four heptad leucine residues play an essential role in mediating Cby homodimerization. Although monomeric Cby is sufficient to bind to beta-catenin and block beta-catenin-mediated transcriptional activation, homodimer formation of Cby is indispensable for its efficient nuclear import.
Figures
Similar articles
-
Structural characterization of partially disordered human Chibby: insights into its function in the Wnt-signaling pathway.Biochemistry. 2011 Feb 8;50(5):715-26. doi: 10.1021/bi101236z. Epub 2011 Jan 14. Biochemistry. 2011. PMID: 21182262 Free PMC article.
-
Nuclear-cytoplasmic shuttling of Chibby controls beta-catenin signaling.Mol Biol Cell. 2010 Jan 15;21(2):311-22. doi: 10.1091/mbc.e09-05-0437. Epub 2009 Nov 25. Mol Biol Cell. 2010. PMID: 19940019 Free PMC article.
-
Chibby cooperates with 14-3-3 to regulate beta-catenin subcellular distribution and signaling activity.J Cell Biol. 2008 Jun 30;181(7):1141-54. doi: 10.1083/jcb.200709091. Epub 2008 Jun 23. J Cell Biol. 2008. PMID: 18573912 Free PMC article.
-
Conformational characterization of the intrinsically disordered protein Chibby: Interplay between structural elements in target recognition.Protein Sci. 2016 Aug;25(8):1420-9. doi: 10.1002/pro.2936. Epub 2016 May 1. Protein Sci. 2016. PMID: 27082063 Free PMC article.
-
Fine-tuning of nuclear-catenin by Chibby and 14-3-3.Cell Cycle. 2009 Jan 15;8(2):210-3. doi: 10.4161/cc.8.2.7394. Epub 2009 Jan 12. Cell Cycle. 2009. PMID: 19158508 Free PMC article. Review.
Cited by
-
Structural Analysis of the 14-3-3ζ/Chibby Interaction Involved in Wnt/β-Catenin Signaling.PLoS One. 2015 Apr 24;10(4):e0123934. doi: 10.1371/journal.pone.0123934. eCollection 2015. PLoS One. 2015. PMID: 25909186 Free PMC article.
-
Structural characterization of partially disordered human Chibby: insights into its function in the Wnt-signaling pathway.Biochemistry. 2011 Feb 8;50(5):715-26. doi: 10.1021/bi101236z. Epub 2011 Jan 14. Biochemistry. 2011. PMID: 21182262 Free PMC article.
-
Structural validation and assessment of AlphaFold2 predictions for centrosomal and centriolar proteins and their complexes.Commun Biol. 2022 Apr 5;5(1):312. doi: 10.1038/s42003-022-03269-0. Commun Biol. 2022. PMID: 35383272 Free PMC article.
-
Chibby functions in Xenopus ciliary assembly, embryonic development, and the regulation of gene expression.Dev Biol. 2014 Nov 15;395(2):287-98. doi: 10.1016/j.ydbio.2014.09.008. Epub 2014 Sep 16. Dev Biol. 2014. PMID: 25220153 Free PMC article.
-
Chibby suppresses growth of human SW480 colon adenocarcinoma cells through inhibition of β-catenin signaling.J Mol Signal. 2012 May 31;7(1):6. doi: 10.1186/1750-2187-7-6. J Mol Signal. 2012. PMID: 22651859 Free PMC article.
References
-
- Wodarz A, Nusse R. Mechanisms of Wnt signaling in development. Annu Rev Cell Dev Biol. 1998;14:59–88. - PubMed
-
- Pinto D, Clevers H. Wnt control of stem cells and differentiation in the intestinal epithelium. Exp Cell Res. 2005;306:357–363. - PubMed
-
- Nusse R. Wnt signaling in disease and in development. Cell Res. 2005;15:28–32. - PubMed
-
- Moon RT, Kohn AD, De Ferrari GV, Kaykas A. WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet. 2004;5:691–701. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
