

Genetic ablation of neural crest cell diversification
- PMID: 19439494
- PMCID: PMC2685722
- DOI: 10.1242/dev.033209
Genetic ablation of neural crest cell diversification
Abstract
The neural crest generates multiple cell types during embryogenesis but the mechanisms regulating neural crest cell diversification are incompletely understood. Previous studies using mutant zebrafish indicated that foxd3 and tfap2a function early and differentially in the development of neural crest sublineages. Here, we show that the simultaneous loss of foxd3 and tfap2a function in zebrafish foxd3(zdf10);tfap2a(low) double mutant embryos globally prevents the specification of developmentally distinct neural crest sublineages. By contrast, neural crest induction occurs independently of foxd3 and tfap2a function. We show that the failure of neural crest cell diversification in double mutants is accompanied by the absence of neural crest sox10 and sox9a/b gene expression, and that forced expression of sox10 and sox9a/b differentially rescues neural crest sublineage specification and derivative differentiation. These results demonstrate the functional necessity for foxd3 and tfap2a for neural crest sublineage specification and that this requirement is mediated by the synergistic regulation of the expression of SoxE family genes. Our results identify a genetic regulatory pathway functionally discrete from the process of neural crest induction that is required for the initiation of neural crest cell diversification during embryonic development.
Figures
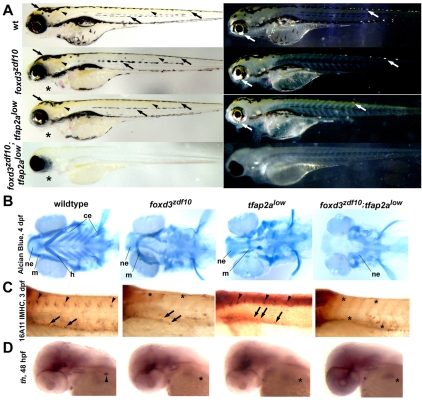
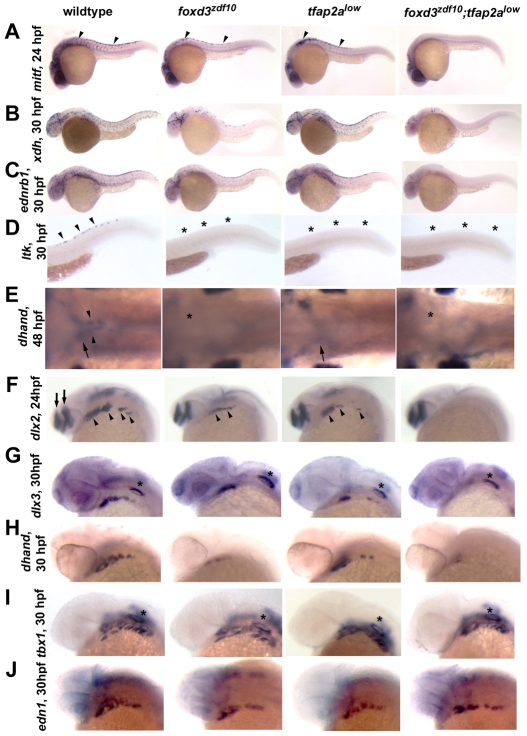
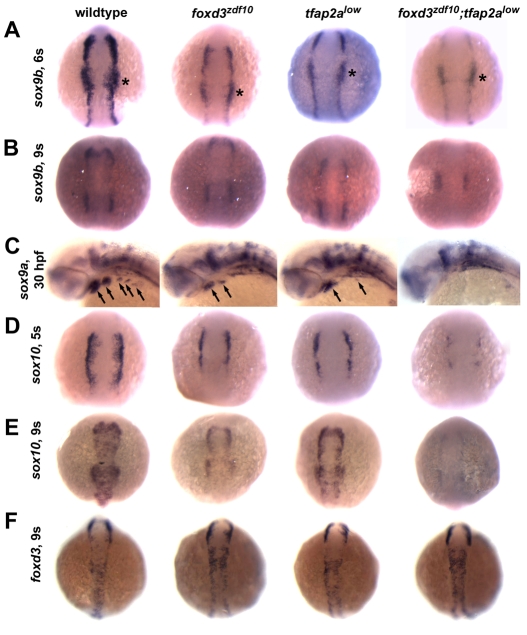
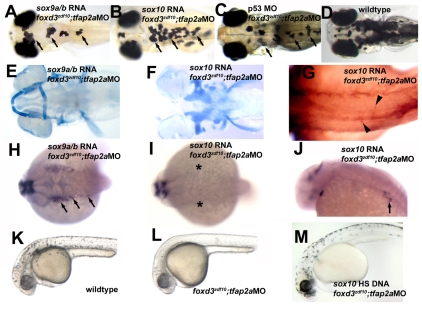
References
-
- Allende, M. L. and Weinberg, E. S. (1994). The expression pattern of two zebrafish achaete-scute homolog (ash) genes is altered in the embryonic brain of the cyclops mutant. Dev. Biol. 166, 509-530. - PubMed
-
- An, M., Luo, R. and Henion, P. D. (2002). Differentiation and maturation of zebrafish dorsal root and sympathetic ganglion neurons. J. Comp. Neurol. 446, 267-275. - PubMed
-
- Barrallo-Gimeno, A., Holzschuh, J., Driever, W. and Knapik, E. W. (2004). Neural crest survival and differentiation in zebrafish depends on mont blanc/tfap2a gene function. Development 131, 1463-1477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
