

Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in Arabidopsis seeds
- PMID: 19439570
- PMCID: PMC2705047
- DOI: 10.1104/pp.109.139352
Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in Arabidopsis seeds
Abstract
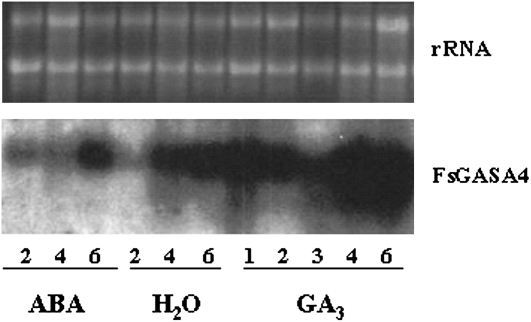
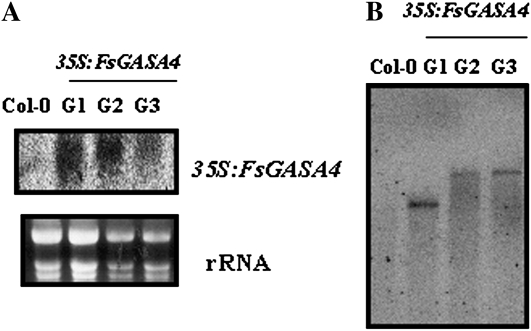
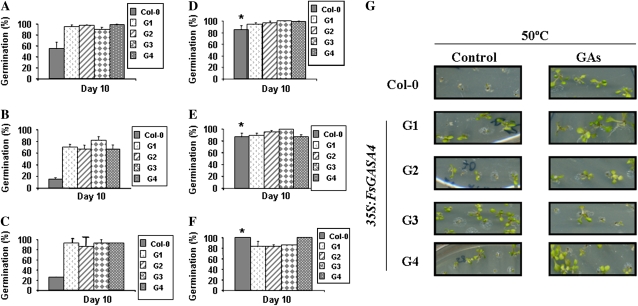
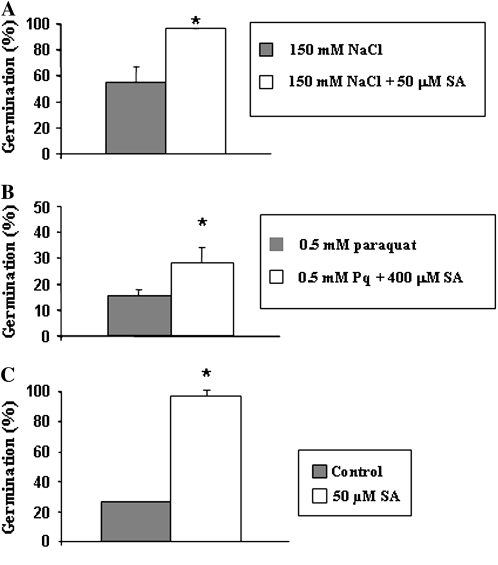
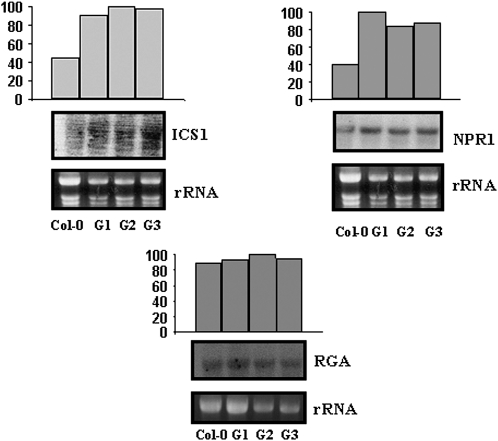
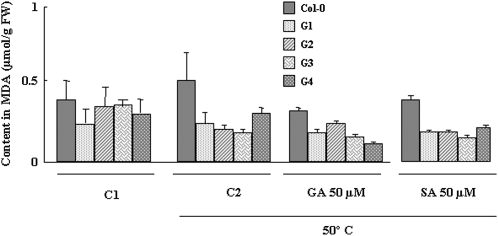
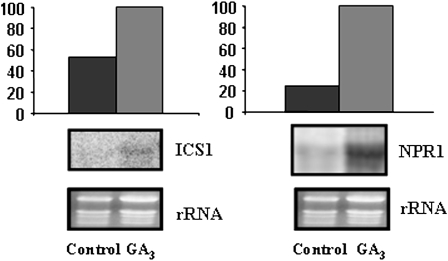
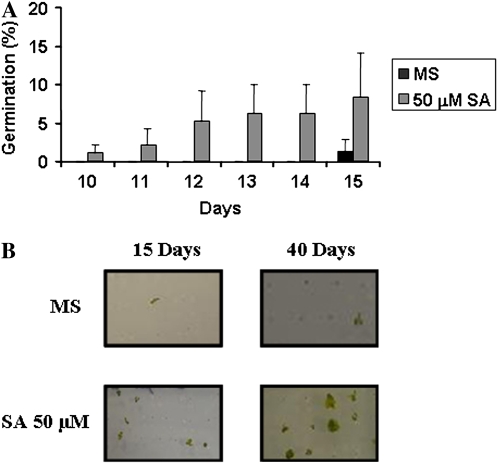
Exogenous application of gibberellic acid (GA(3)) was able to reverse the inhibitory effect of salt, oxidative, and heat stresses in the germination and seedling establishment of Arabidopsis (Arabidopsis thaliana), this effect being accompanied by an increase in salicylic acid (SA) levels, a hormone that in recent years has been implicated in plant responses to abiotic stress. Furthermore, this treatment induced an increase in the expression levels of the isochorismate synthase1 and nonexpressor of PR1 genes, involved in SA biosynthesis and action, respectively. In addition, we proved that transgenic plants overexpressing a gibberellin (GA)-responsive gene from beechnut (Fagus sylvatica), coding for a member of the GA(3) stimulated in Arabidopsis (GASA) family (FsGASA4), showed a reduced GA dependence for growth and improved responses to salt, oxidative, and heat stress at the level of seed germination and seedling establishment. In 35S:FsGASA4 seeds, the improved behavior under abiotic stress was accompanied by an increase in SA endogenous levels. All these data taken together suggest that this GA-responsive gene and exogenous addition of GAs are able to counteract the inhibitory effects of these adverse environmental conditions in seed germination and seedling growth through modulation of SA biosynthesis. Furthermore, this hypothesis is supported by the fact that sid2 mutants, impaired in SA biosynthesis, are more sensitive to salt stress than wild type and are not affected by exogenous application of GA(3).
Figures

Comment in
-
Cross-talk between gibberellins and salicylic acid in early stress responses in Arabidopsis thaliana seeds.Plant Signal Behav. 2009 Aug;4(8):750-1. doi: 10.1104/pp.109.139352. Epub 2009 Aug 2. Plant Signal Behav. 2009. PMID: 19820299 Free PMC article.
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van der Straeten D, Peng JR, Harberd NP (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311 91–94 - PubMed
-
- Ananieva EA, Alexieva VS, Popova LP (2002) Treatment with salicylic acid decreases the effects of paraquat on photosynthesis. J Plant Physiol 159 685–693
-
- Ananieva EA, Christov KN, Popova LP (2004) Exogenous treatment with salicylic acid leads to increased antioxidant capacity in leaves of barley plants exposed to paraquat. J Plant Physiol 161 319–328 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
