

Nck proteins maintain the adult glomerular filtration barrier
- PMID: 19443634
- PMCID: PMC2709686
- DOI: 10.1681/ASN.2009010056
Nck proteins maintain the adult glomerular filtration barrier
Abstract
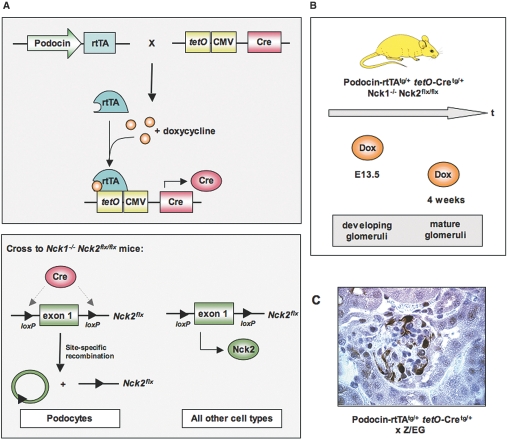
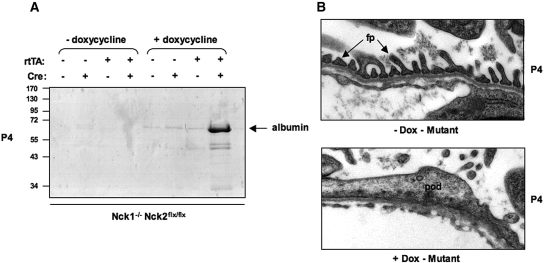
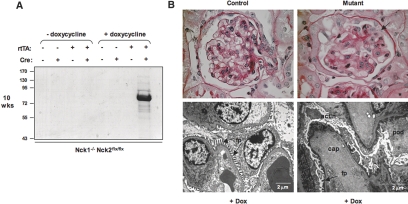
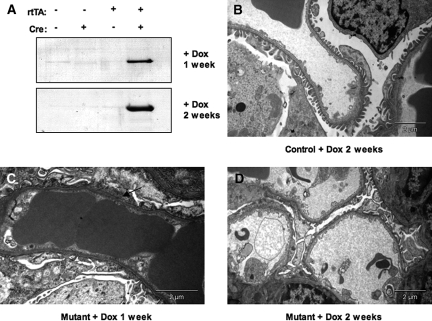
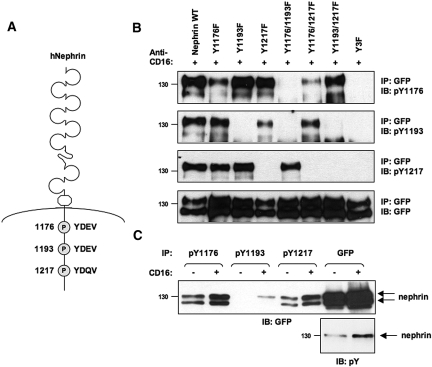
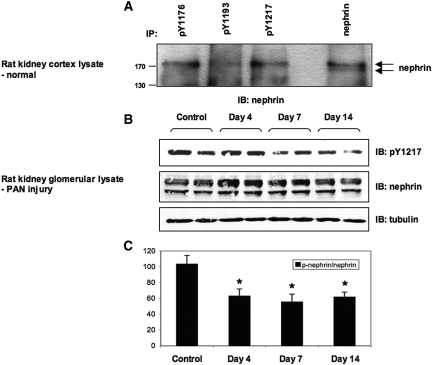
Within the glomerulus, the scaffolding protein nephrin bridges the actin-rich foot processes that extend from adjacent podocytes to form the slit diaphragm. Mutations affecting a number of slit diaphragm proteins, including nephrin, cause glomerular disease through rearrangement of the actin cytoskeleton and disruption of the filtration barrier. We recently established that the Nck family of Src homology 2 (SH2)/SH3 cytoskeletal adaptor proteins can mediate nephrin-dependent actin reorganization. Formation of foot processes requires expression of Nck in developing podocytes, but it is unknown whether Nck maintains podocyte structure and function throughout life. Here, we used an inducible transgenic strategy to delete Nck expression in adult mouse podocytes and found that loss of Nck expression rapidly led to proteinuria, glomerulosclerosis, and altered morphology of foot processes. We also found that podocyte injury reduced phosphorylation of nephrin in adult kidneys. These data suggest that Nck is required to maintain adult podocytes and that phosphotyrosine-based interactions with nephrin may occur in foot processes of resting, mature podocytes.
Figures
Comment in
-
The promise of well-being: stay in shape with N(i)ck.J Am Soc Nephrol. 2009 Jul;20(7):1425-7. doi: 10.1681/ASN.2009040453. Epub 2009 Jun 4. J Am Soc Nephrol. 2009. PMID: 19497966 No abstract available.
References
-
- Pavenstadt H, Kriz W, Kretzler M: Cell biology of the glomerular podocyte. Physiol Rev 83: 253–307, 2003 - PubMed
-
- Michaud JL, Kennedy CR: The podocyte in health and disease: Insights from the mouse. Clin Sci (Lond) 112: 325–335, 2007 - PubMed
-
- Wiggins RC: The spectrum of podocytopathies: A unifying view of glomerular diseases. Kidney Int 71: 1205–1214, 2007 - PubMed
-
- Patrakka J, Tryggvason K: Nephrin—A unique structural and signalling protein of the kidney filter. Trends Mol Med 13: 396–403, 2007 - PubMed
-
- Lahdenpera J, Kilpelainen P, Liu XL, Pikkarainen T, Reponen P, Ruotsalainen V, Tryggvason K: Clustering-induced tyrosine phosphorylation of nephrin by Src family kinases. Kidney Int 64: 404–413, 2003 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
