

LKB1 regulates polarity remodeling and adherens junction formation in the Drosophila eye
- PMID: 19443685
- PMCID: PMC2690039
- DOI: 10.1073/pnas.0812469106
LKB1 regulates polarity remodeling and adherens junction formation in the Drosophila eye
Abstract
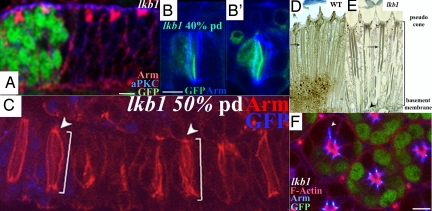
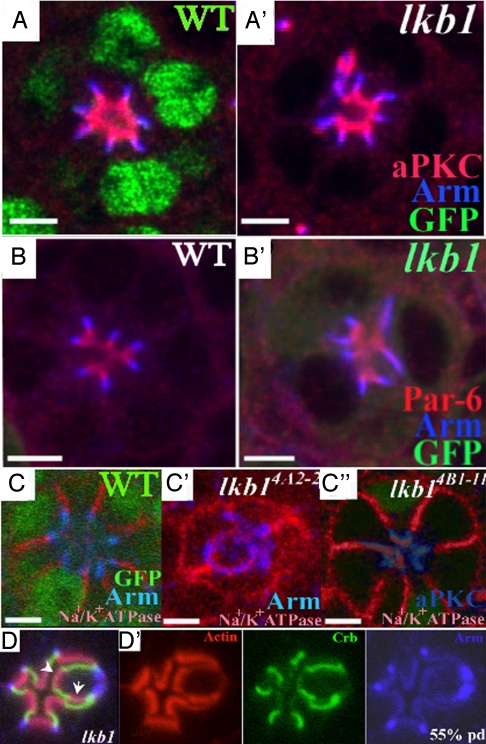
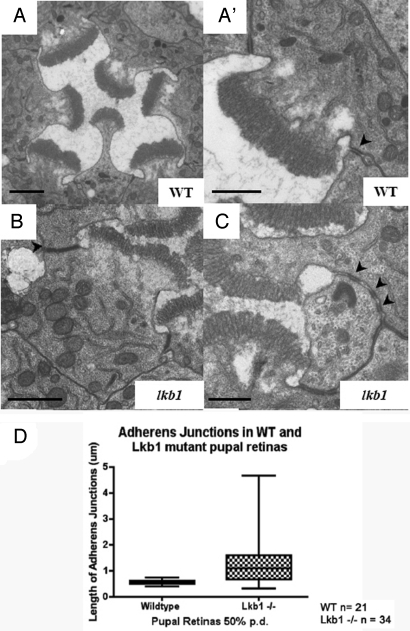
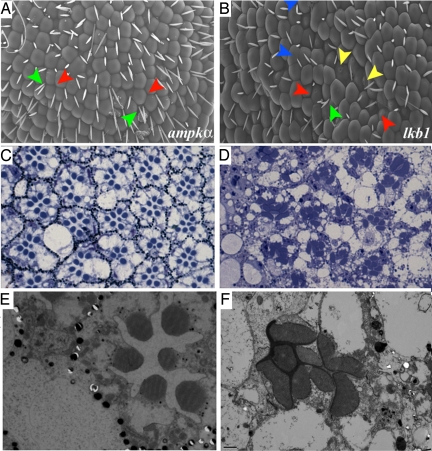
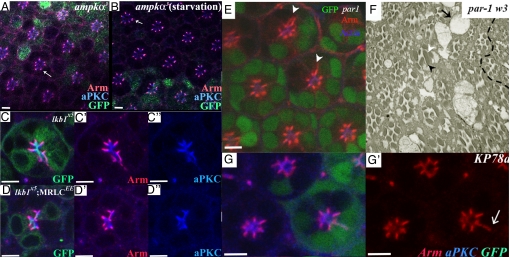
The serine-threonine kinase LKB1 regulates cell polarity from Caenorhabditis elegans to man. Loss of lkb1 leads to a cancer predisposition, known as Peutz-Jeghers Syndrome. Biochemical analysis indicates that LKB1 can phosphorylate and activate a family of AMPK- like kinases, however, the precise contribution of these kinases to the establishment and maintenance of cell polarity is still unclear. Recent studies propose that LKB1 acts primarily through the AMP kinase to establish and/or maintain cell polarity. To determine whether this simple model of how LKB1 regulates cell polarity has relevance to complex tissues, we examined lkb1 mutants in the Drosophila eye. We show that adherens junctions expand and apical, junctional, and basolateral domains mix in lkb1 mutants. Surprisingly, we find LKB1 does not act primarily through AMPK to regulate cell polarity in the retina. Unlike lkb1 mutants, ampk retinas do not show elongated rhabdomeres or expansion of apical and junctional markers into the basolateral domain. In addition, nutrient deprivation does not reveal a more dramatic polarity phenotype in lkb1 photoreceptors. These data suggest that AMPK is not the primary target of LKB1 during eye development. Instead, we find that a number of other AMPK-like kinase, such as SIK, NUAK, Par-1, KP78a, and KP78b show phenotypes similar to weak lkb1 loss of function in the eye. These data suggest that in complex tissues, LKB1 acts on an array of targets to regulate cell polarity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Watts JL, Morton DG, Bestman J, Kemphues KJ. The C. elegans par-4 gene encodes a putative serine–threonine kinase required for establishing embryonic asymmetry. Development. 2000;127:1467–1475. - PubMed
-
- Martin SG, St Johnston D. A role for Drosophila LKB1 in anterior–posterior axis formation and epithelial polarity. Nature. 2003;421:379–384. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
