

Review
doi: 10.1038/nri2550.
Autoimmune T cell responses in the central nervous system
Affiliations
- PMID: 19444307
- PMCID: PMC2813731
- DOI: 10.1038/nri2550
Item in Clipboard
Review
Autoimmune T cell responses in the central nervous system
Nat Rev Immunol.
2009 Jun.
Abstract
Autoreactive T cell responses have a crucial role in central nervous system (CNS) diseases such as multiple sclerosis. Recent data indicate that CNS autoimmunity can be mediated by two distinct lineages of CD4+ T cells that are defined by the production of either interferon-gamma or interleukin-17. The activity of these CD4+ T cell subsets within the CNS influences the pathology and clinical course of disease. New animal models show that myelin-specific CD8+ T cells can also mediate CNS autoimmunity. This Review focuses on recent progress in delineating the pathogenic mechanisms, regulation and interplay between these different T cell subsets in CNS autoimmunity.
Figures
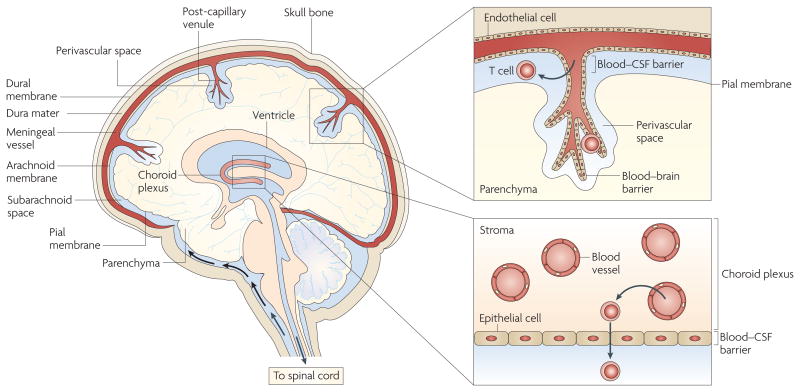
Activated T cells can enter the subarachnoid space by migrating from blood vessels into the stroma of the choroid plexus and then crossing the blood–cerebrospinal fluid (CSF) barrier surrounding the choroid plexus stroma, which comprises epithelial cells joined by tight junctions. Activated T cells can also enter the subarachnoid space by extravasating through the cell wall of meningeal venules, which consists of endothelial cells connected by tight junctions. In addition, activated T cells can cross the blood–brain barrier surrounding post-capillary venules that penetrate the brain parenchyma, which comprises endothelial cells connected by tight junctions. T cells crossing the blood–brain barrier enter the perivascular space, which is the region between the basement membrane connected to the blood vessel endothelial cells and the glial limitans (which is composed of astrocyte feet and microglial cell; not shown).
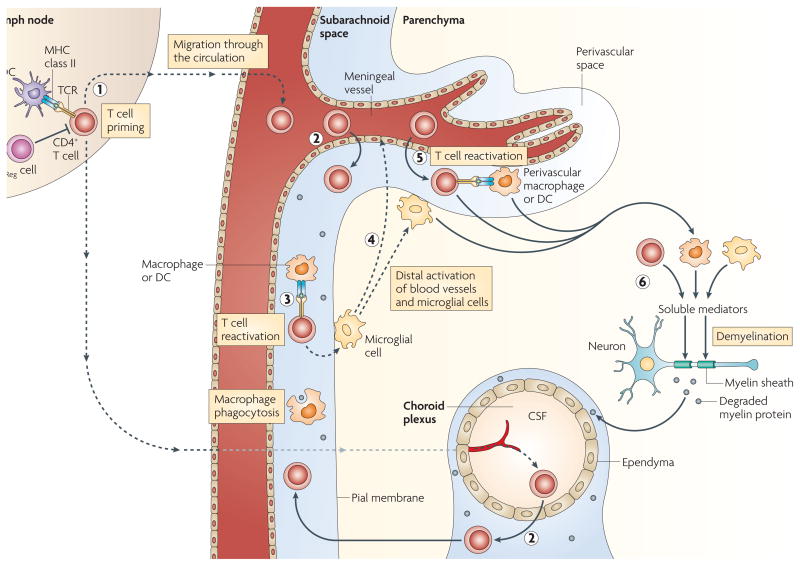
CD4+ T cells are primed in the periphery by dendritic cells (DCs) presenting myelin (or myelin crossreactive) epitopes. Antigen-presenting cells (APCs) residing in the central nervous system (CNS) can capture myelin antigens in situ and migrate to the cervical lymph nodes. Alternatively, soluble myelin antigens can drain from the CNS to lymph nodes to be phagocytosed by local APCs (1). CD4+ T cells enter the subarachnoid space by crossing the blood–cerebrospinal fluid (CSF) barrier either in the choroid plexus or the meningeal venules (2); the T cells are re-activated within the subarachnoid space by MHC class II-expressing macrophages and DCs expressing myelin epitopes (3). Reactivated T cells activate microglial cells in the subpial region, triggering activation of distal microglial cells and blood vessels (4). Activated T cells adhere to and cross the activated blood–brain barrier, enter the perivascular space and are reactivated by perivascular macrophages and DCs (5). T cells enter the parenchyma and, together with activated macrophages and microglial cells, secrete soluble mediators that trigger demyelination (6). TReg, regulatory T.
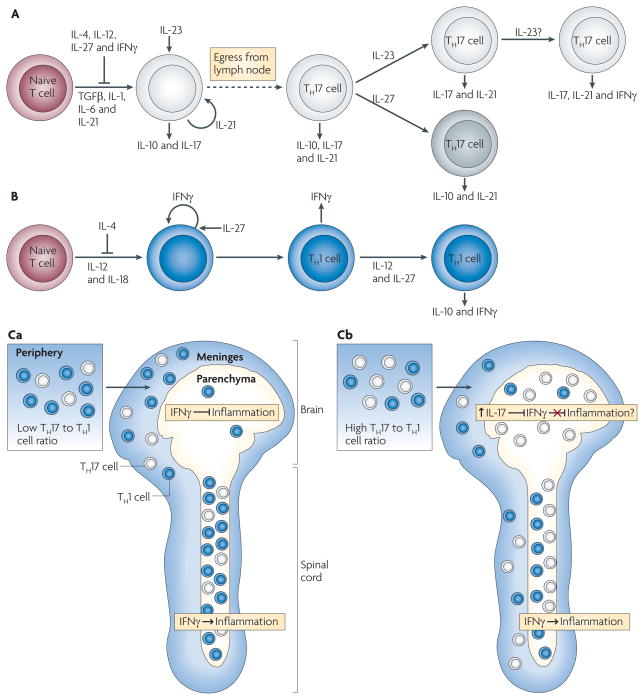
A. | Naive T cells differentiate in the presence of transforming growth factor-β (TGFβ), interleukin-1 (IL-1), IL-6 and IL-21 into an early T helper 17 (TH17) cell stage that also produces IL-10 and IL-17. IL-4, IL-12, IL-27 and interferon-γ (IFNγ) inhibit TH17 cell differentiation, and IL-21 acts in an autocrine manner at this stage. IL-23 promotes TH17 cell differentiation, decreases IL-10 production and stabilizes the IL-17-producing phenotype. IL-23 may facilitate co-expression of IFNγ under some circumstances. IL-27 can suppress IL-17 and induce IL-10 production by TH17 cells. B | Differentiation of TH1 cells occurs in the presence of IL-12 and IL-18, with IFNγ acting in an autocrine manner. TH1 cell differentiation is inhibited by IL-4, and IL-27 can enhance IFNγ production by differentiating TH1 cells. Exposure to IL-12 and IL-27 induces differentiated TH1 cells to produce IL-10 and IFNγ. Differentiated TH17 and TH1 cells express other pro-inflammatory cytokines (not depicted). Ca | Myelin-specific T cells with a low TH17 to TH1 cell ratio are primed in the periphery and infiltrate the meninges of both the brain and spinal cord. The T cells enter the spinal cord parenchyma and induce inflammation in an IFNγ-mediated signalling-dependent manner. The few T cells that enter the brain parenchyma, where IFNγ-mediated signalling inhibits inflammation, do not recruit inflammatory cells. Cb | Myelin-specific T cells primed in the periphery with a high TH17 to TH1 cell ratio infiltrate the meninges and parenchyma of both the brain and spinal cord. The higher TH17 to TH1 cell ratio in the brain is associated with a disproportionate increase in IL-17 production compared with the spinal cord, which may overcome the inhibitory IFNγ-mediated signalling in the brain parenchyma.
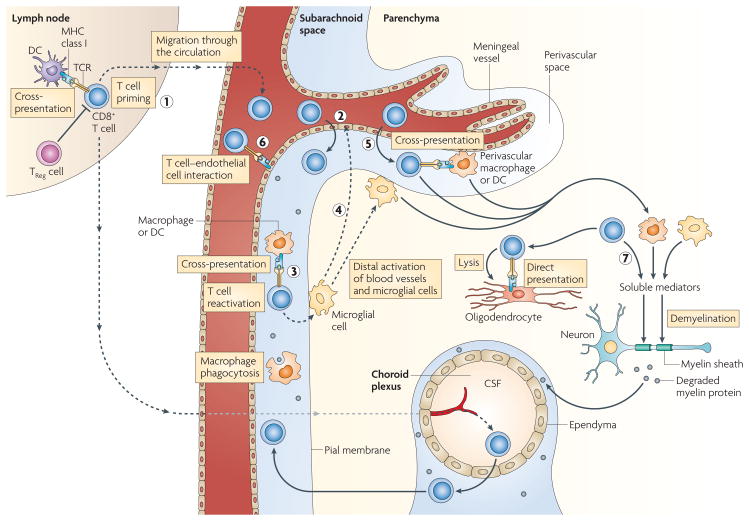
CD8+ T cells are primed by cross-presentation by dendritic cells (DCs) in the lymph nodes (1) and follow the same steps (2–5) as CD4+ T cells (see FIG. 2). However, CD8+ T cells are activated by macrophages, microglial cells and DCs only by cross-presentation; endothelial cells can directly present antigen if they have access to myelin epitopes (6). Activated macrophages, microglial cells and CD8+ T cells secrete soluble mediators, and CD8+ T cells can directly lyse oligodendrocytes expressing MHC class I and myelin epitopes (7). CSF, cerebrospinal fluid; TCR, T cell receptor; TReg, regulator T.
References
-
- McFarland HF, Martin R. Multiple sclerosis: a complicated picture of autoimmunity. Nature Immunol. 2007;8:913–919. - PubMed
-
- Lucchinetti C, et al. Heterogeneity of multiple sclerosis lesions: implications for the pathogenesis of demyelination. Ann Neurol. 2000;47:707–717. - PubMed
-
- Raine C. In: Multiple Sclerosis: Clinical and Pathogenetic Basis. Raine CS, McFarland HF, Tourtellotte WW, editors. Chapman and Hall; London: 1997. pp. 243–286.
-
- Ransohoff RM, Kivisakk P, Kidd G. Three or more routes for leukocyte migration into the central nervous system. Nature Rev Immunol. 2003;3:569–581. - PubMed
-
- Huseby ES, Sather B, Huseby PG, Goverman J. Age-dependent T cell tolerance and autoimmunity to myelin basic protein. Immunity. 2001;14:471–481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
