

Deletion of steroid 5 alpha-reductase 2 gene in male pseudohermaphroditism
- PMID: 1944596
- PMCID: PMC4451825
- DOI: 10.1038/354159a0
Deletion of steroid 5 alpha-reductase 2 gene in male pseudohermaphroditism
Abstract
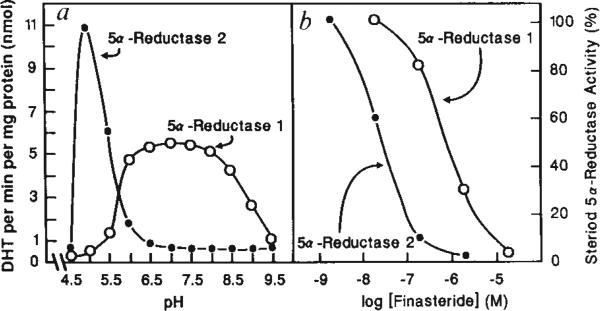
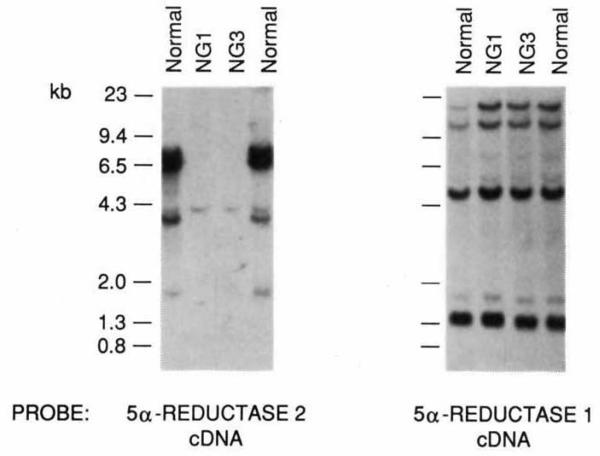
The conversion of testosterone into dihydrotestosterone by steroid 5 alpha-reductase is a key reaction in androgen action, and is essential both for the formation of the male phenotype during embryogenesis and for androgen-mediated growth of tissues such as the prostate. Single gene defects that impair this conversion lead to pseudohermaphroditism in which 46X,Y males have male internal urogenital tracts, but female external genitalia. We have described the isolation of a human 5 alpha-reductase complementary DNA from prostate. Subsequent cloning and genetic studies showed that this gene (designated 5 alpha-reductase 1) was normal in patients with 5 alpha-reductase deficiency. We report here the isolation of a second 5 alpha-reductase cDNA by expression cloning and the polymerase chain reaction. The biochemical and pharmacological properties of this cDNA-encoded enzyme (designated 5 alpha-reductase 2) are consistent with it being the major isozyme in genital tissue. A deletion in this gene is present in two related individuals with male pseudohermaphroditism caused by 5 alpha-reductase deficiency. These results verify the existence of at least two 5 alpha-reductases in man and provide insight into a fundamental hormone-mediated event in male sexual differentiation.
Figures


References
-
- Wilson. J. D. Harvey Led. 1985;79:145–172. - PubMed
-
- Wilson JD. Endocrinology. 1975;25:491–508.
-
- Griffin JE, Wilson JD. In: The Metabolic Basis of Inherited Disease. Scriver CR, Beaudet AL, Sly WS, Valle D, editors. McGraw-Hill; New York: 1989. pp. 1919–1944.
-
- Andersson S, Davis D, Dahlback, Jornvail H, Russell DW. J. biol. Chem. 1989;264:8222–8229. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
