

Large conductance calcium-activated potassium channels affect both spontaneous firing and intracellular calcium concentration in cerebellar Purkinje neurons
- PMID: 19446607
- PMCID: PMC2723190
- DOI: 10.1016/j.neuroscience.2009.05.016
Large conductance calcium-activated potassium channels affect both spontaneous firing and intracellular calcium concentration in cerebellar Purkinje neurons
Abstract
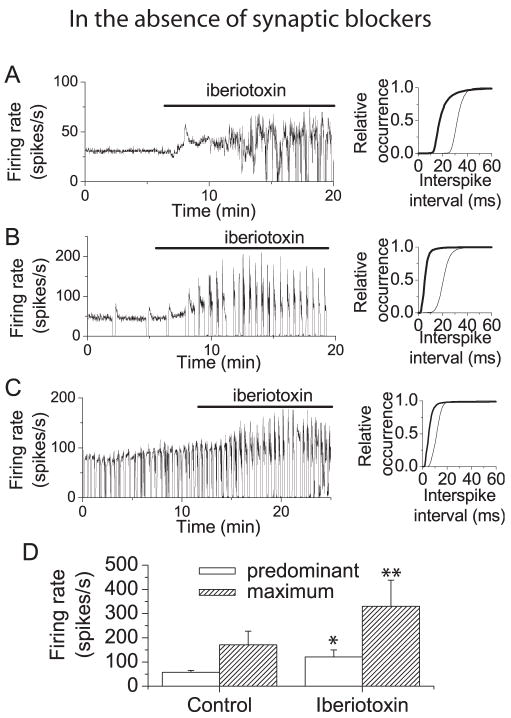
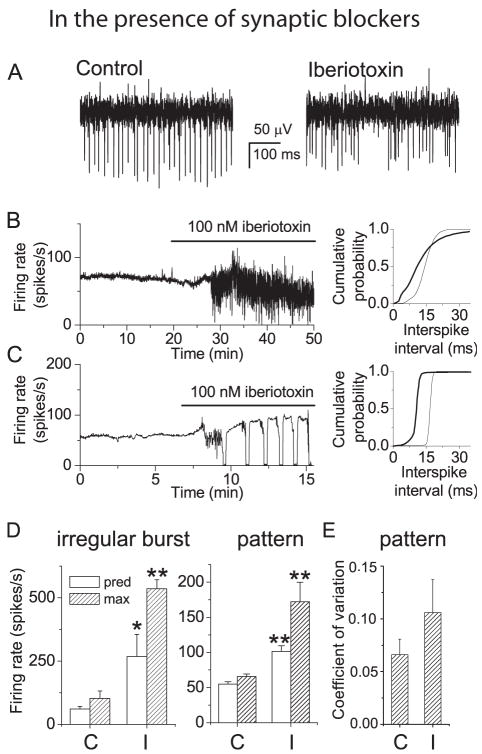
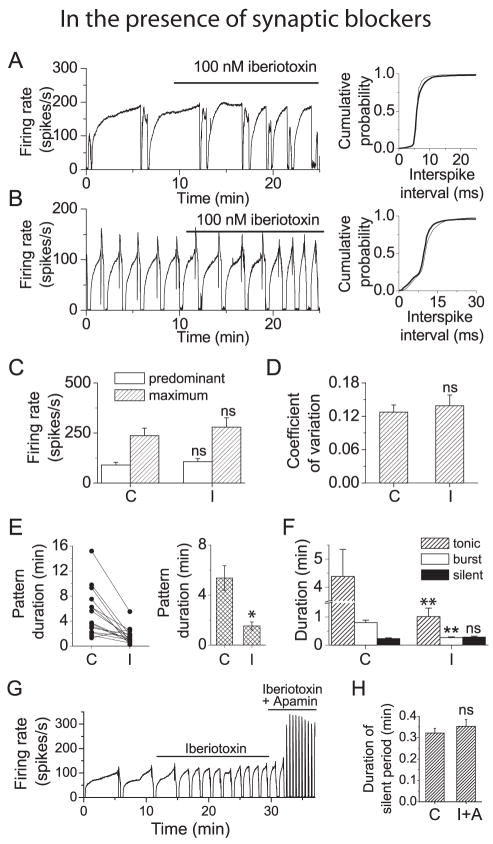
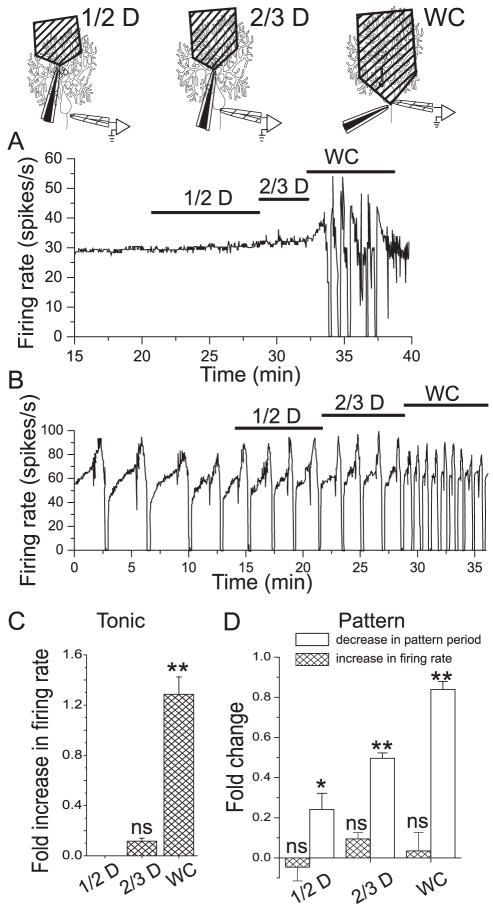
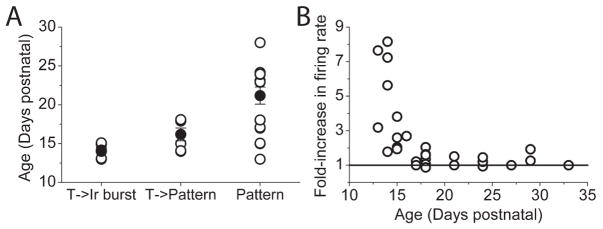
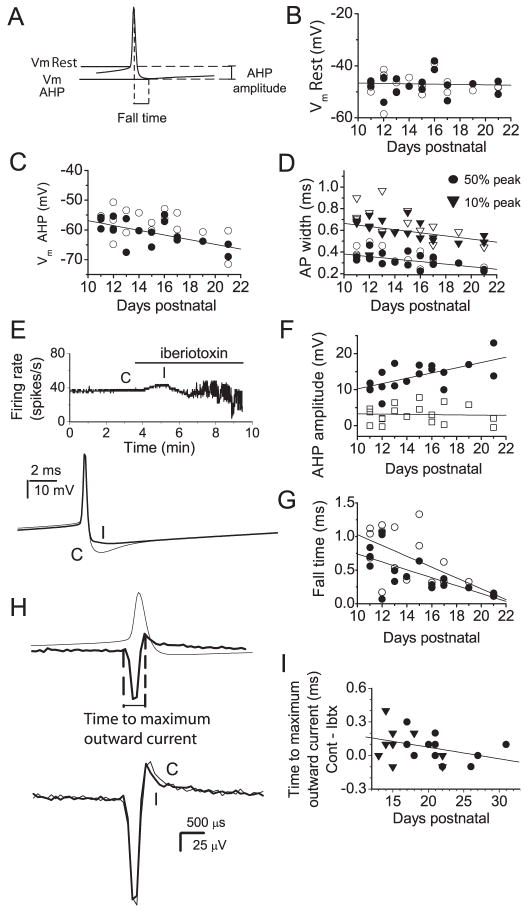
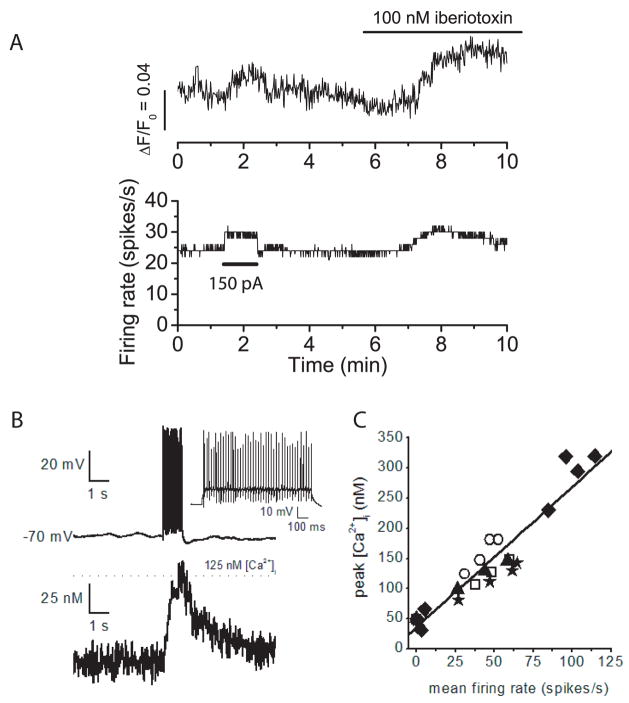
We investigated the contribution of large conductance calcium-activated potassium (BK) channels to spontaneous activity of cerebellar Purkinje neurons in mice and rats. In Purkinje neurons which fire tonically, block of BK channels increased the firing rate and caused the neurons to fire irregularly. In Purkinje neurons which exhibited a trimodal pattern of activity, present primarily in mature animals, block of BK channels had little effect on firing rate or regularity but shortened the single cycle duration of the trimodal pattern. The contribution of BK channels to the action potential waveform was also examined. BK channels contributed a brief afterhyperpolarization (AHP) of approximately 3 mV which followed each action potential, but made little contribution to action potential repolarization. The amplitude of the BK-dependent AHP did not change with age although there was an increase in the total AHP. The difference in the contribution of BK channels to the firing rate among the two populations of Purkinje neurons was the consequence of the decrease in the fractional contribution of BK channels to the AHP. We also found that block of BK channels increases intracellular calcium concentration during spontaneous firing. Thus, although BK channels do not affect action potential repolarization, they nevertheless control calcium entry with each action potential by contributing to the AHP.
Figures
References
-
- Giangiacomo KM, Garcia ML, McManus OB. Mechanism of iberiotoxin block of the large-conductance calcium- activated potassium channel from bovine aortic smooth muscle. Biochemistry. 1992;31:6719–6727. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
