

Bcl-2 blocks 2-methoxyestradiol induced leukemia cell apoptosis by a p27(Kip1)-dependent G1/S cell cycle arrest in conjunction with NF-kappaB activation
- PMID: 19447221
- PMCID: PMC2782607
- DOI: 10.1016/j.bcp.2009.03.017
Bcl-2 blocks 2-methoxyestradiol induced leukemia cell apoptosis by a p27(Kip1)-dependent G1/S cell cycle arrest in conjunction with NF-kappaB activation
Abstract
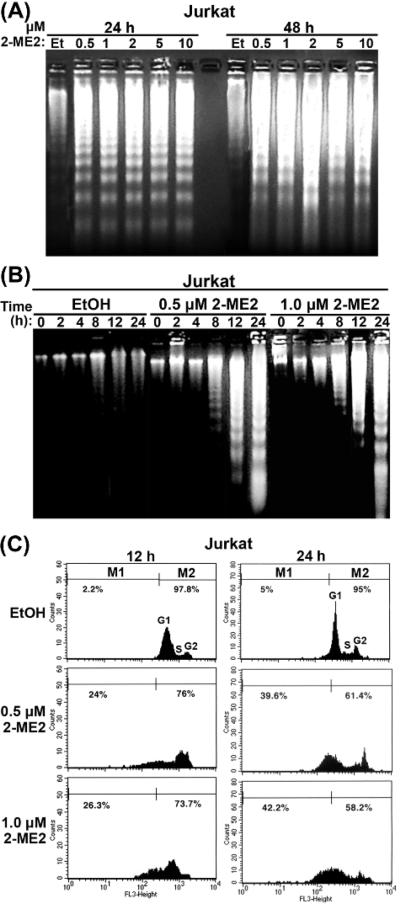
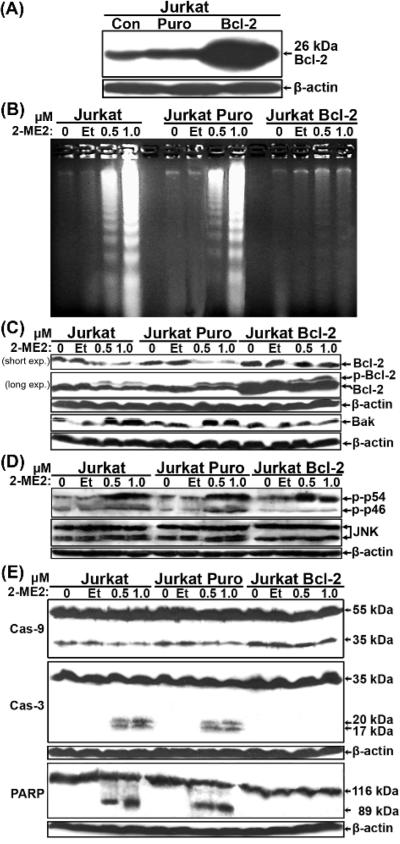
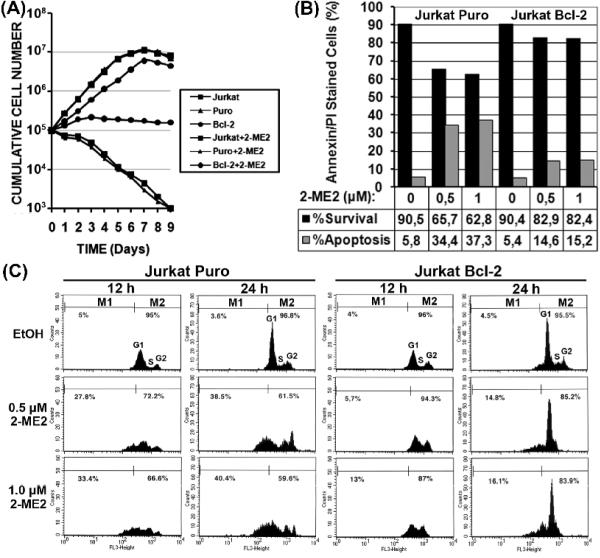
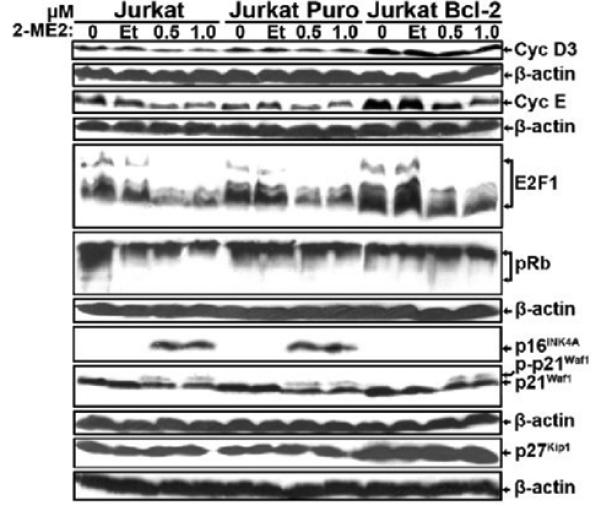
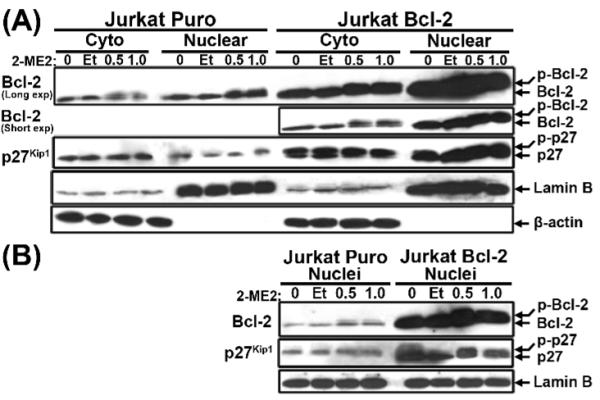
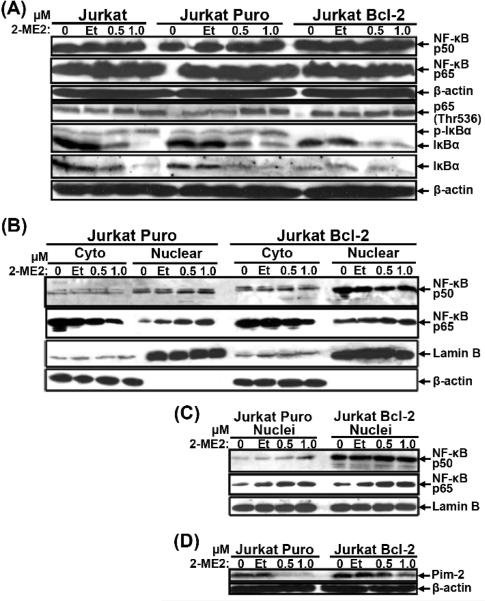
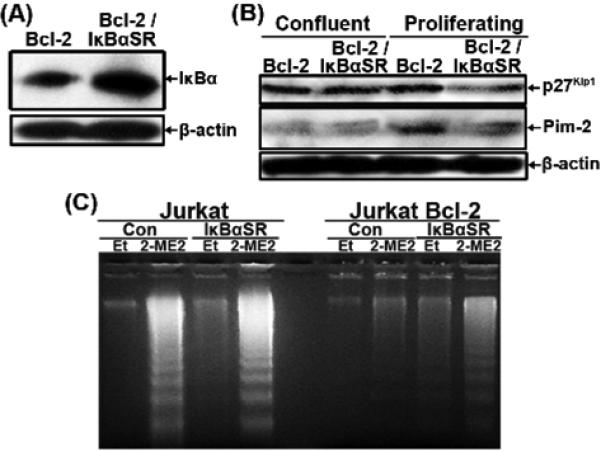
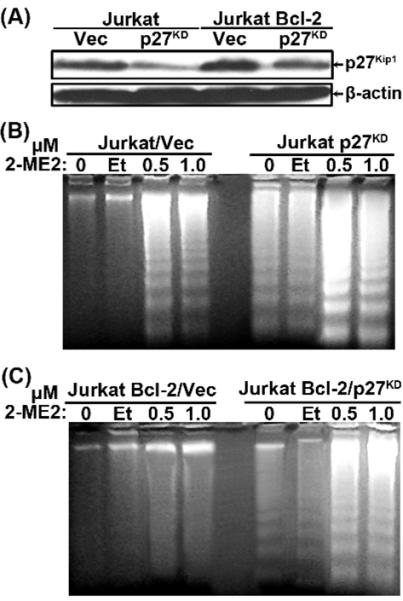
2-Methoxyestradiol (2-ME2) induces leukemia cells to undergo apoptosis in association with Bcl-2 inactivation but the mechanisms whereby Bcl-2 contributes to protection against programmed cell death in this context remain unclear. Here we showed that 2-ME2 inhibited the proliferation of Jurkat leukemia cells by markedly suppressing the levels of cyclins D3 and E, E2F1 and p21(Cip1/Waf1) and up-regulating p16(INK4A). Further, 2-ME2 induced apoptosis of Jurkat cells in association with down-regulation and phosphorylation of Bcl-2 (as mediated by JNK), up-regulation of Bak, activation of caspases-9 and -3 and PARP-1 cleavage. To determine the importance and mechanistic role of Bcl-2 in this process, we enforced its expression in Jurkat cells by retroviral transduction. Enforcing Bcl-2 expression in Jurkat cells abolished 2-ME2-induced apoptosis and instead produced a G1/S phase cell cycle arrest in association with markedly increased levels of p27(Kip1). Bcl-2 and p27(Kip1) were localized mainly in the nucleus in these apoptotic resistant cells. Interestingly, NF-kappaB activity and p50 levels were increased by 2-ME2 and suppression of NF-kappaB signaling reduced p27(Kip1) expression and sensitized cells to 2-ME2-induced apoptosis. Importantly, knocking-down p27(Kip1) in Jurkat Bcl-2 cells sensitized them to spontaneous and 2-ME2-induced apoptosis. Thus, Bcl-2 prevented the 2-ME2-induced apoptotic response by orchestrating a p27(Kip1)-dependent G1/S phase arrest in conjunction with activating NF-kappaB. Thus, we achieved a much better understanding of the penetrance and mechanistic complexity of Bcl-2 dependent anti-apoptotic pathways in cancer cells and why Bcl-2 inactivation is so critical for the efficacy of apoptosis and anti-proliferative inducing drugs like 2-ME2.
Figures
References
-
- Fotsis T, Zhang Y, Pepper MS, Adlercreutz H, Montesano R, Nawroth PP, et al. The endogenous estrogen metabolite 2-methoxyoestradiol inhibits angiogenesis and suppresses tumour growth. Nature. 1994;368:237–39. - PubMed
-
- Klauber N, Parangi S, Flynn E, Hamel E, D’Amato RJ. Inhibition of angiogenesis and breast cancer in mice by microtubule inhibitors 2-methoxyestradiol and taxol. Cancer Res. 1997;57:81–86. - PubMed
-
- Pribluda VS, Gubish ER, Jr, LaVallee TM, Treston A, Swartz GM, Green SJ. 2-methoxyestradiol: an endogenous antiangiogenic and antiproliferative drug candidate. Cancer Metastasis Rev. 2000;19:173–79. - PubMed
-
- Ryoo JJ, Cole CE, Anderson KC. Novel therapies for multiple myeloma. Blood Rev. 2002;16:167–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
