

Molecular physiology and genetics of Na+-independent SLC4 anion exchangers
- PMID: 19448077
- PMCID: PMC2683012
- DOI: 10.1242/jeb.029454
Molecular physiology and genetics of Na+-independent SLC4 anion exchangers
Abstract
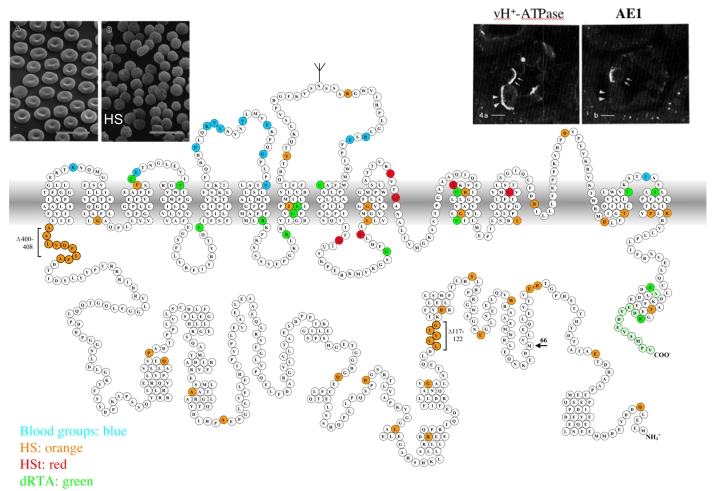
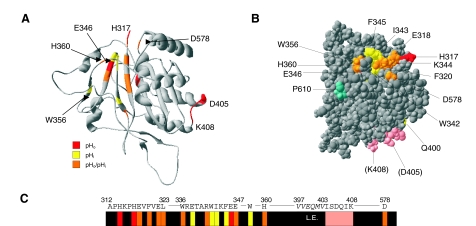
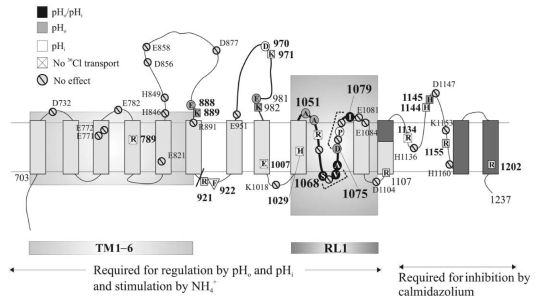
Plasmalemmal Cl(-)/HCO(3)(-) exchangers are encoded by the SLC4 and SLC26 gene superfamilies, and function to regulate intracellular pH, [Cl(-)] and cell volume. The Cl(-)/HCO(3)(-) exchangers of polarized epithelial cells also contribute to transepithelial secretion and reabsorption of acid-base equivalents and Cl(-). This review focuses on Na(+)-independent electroneutral Cl(-)/HCO(3)(-) exchangers of the SLC4 family. Human SLC4A1/AE1 mutations cause the familial erythroid disorders of spherocytic anemia, stomatocytic anemia and ovalocytosis. A largely discrete set of AE1 mutations causes familial distal renal tubular acidosis. The Slc4a2/Ae2(-/-) mouse dies before weaning with achlorhydria and osteopetrosis. A hypomorphic Ae2(-/-) mouse survives to exhibit male infertility with defective spermatogenesis and a syndrome resembling primary biliary cirrhosis. A human SLC4A3/AE3 polymorphism is associated with seizure disorder, and the Ae3(-/-) mouse has increased seizure susceptibility. The transport mechanism of mammalian SLC4/AE polypeptides is that of electroneutral Cl(-)/anion exchange, but trout erythroid Ae1 also mediates Cl(-) conductance. Erythroid Ae1 may mediate the DIDS-sensitive Cl(-) conductance of mammalian erythrocytes, and, with a single missense mutation, can mediate electrogenic SO(4)(2-)/Cl(-) exchange. AE1 trafficking in polarized cells is regulated by phosphorylation and by interaction with other proteins. AE2 exhibits isoform-specific patterns of acute inhibition by acidic intracellular pH and independently by acidic extracellular pH. In contrast, AE2 is activated by hypertonicity and, in a pH-independent manner, by ammonium and by hypertonicity. A growing body of structure-function and interaction data, together with emerging information about physiological function and structure, is advancing our understanding of SLC4 anion exchangers.
Figures

References
-
- Adair-Kirk, T. L., Dorsey, F.C. and Cox, J. V. (2003). Multiple cytoplasmic signals direct the intracellular trafficking of chicken kidney AE1 anion exchangers in MDCK cells. J. Cell Sci. 116, 655-663. - PubMed
-
- Alper, S. L. (2002). Genetic diseases of acid-base transporters. Annu. Rev. Physiol. 64, 899-923. - PubMed
-
- Alper, S. L. (2006). Molecular physiology of SLC4 anion exchangers. Exp. Physiol. 91, 153-161. - PubMed
-
- Alper, S. L., Kopito, R. R., Libresco, S. M. and Lodish, H. F. (1988). Cloning and characterization of a murine band 3-related cDNA from kidney and from a lymphoid cell line. J. Biol. Chem. 263, 17092-17099. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
