

The cation diffusion facilitator gene cdf-2 mediates zinc metabolism in Caenorhabditis elegans
- PMID: 19448268
- PMCID: PMC2728845
- DOI: 10.1534/genetics.109.103614
The cation diffusion facilitator gene cdf-2 mediates zinc metabolism in Caenorhabditis elegans
Abstract
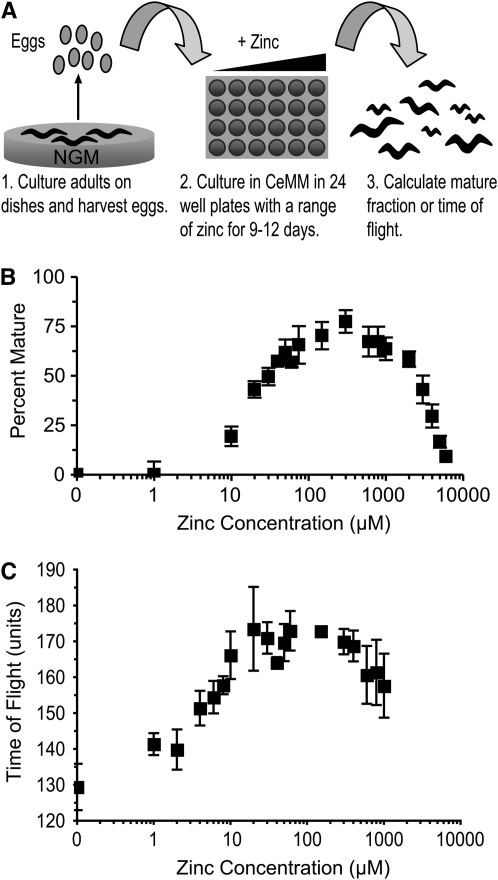
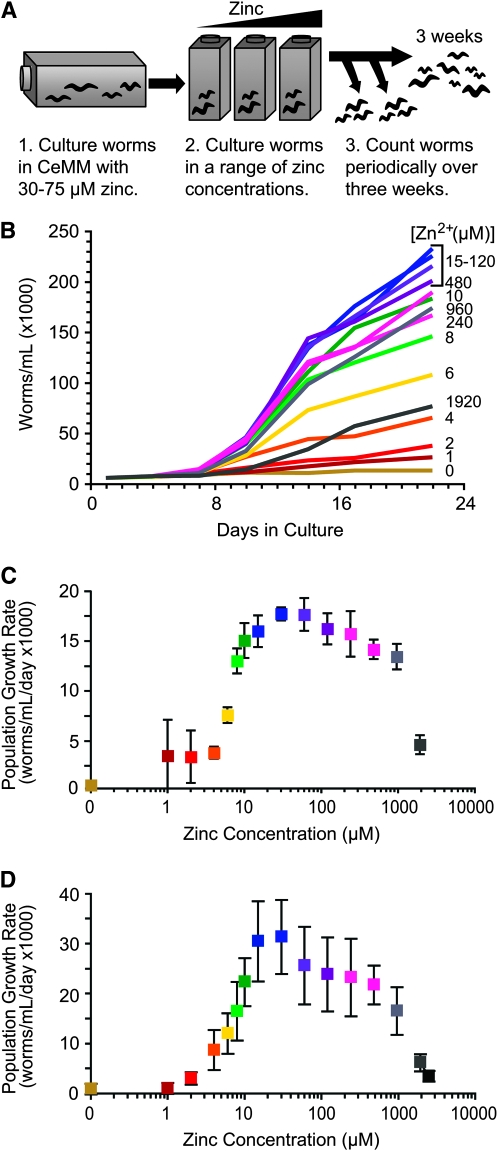
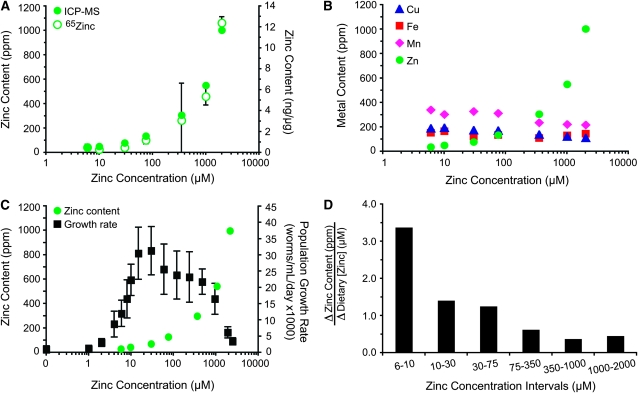
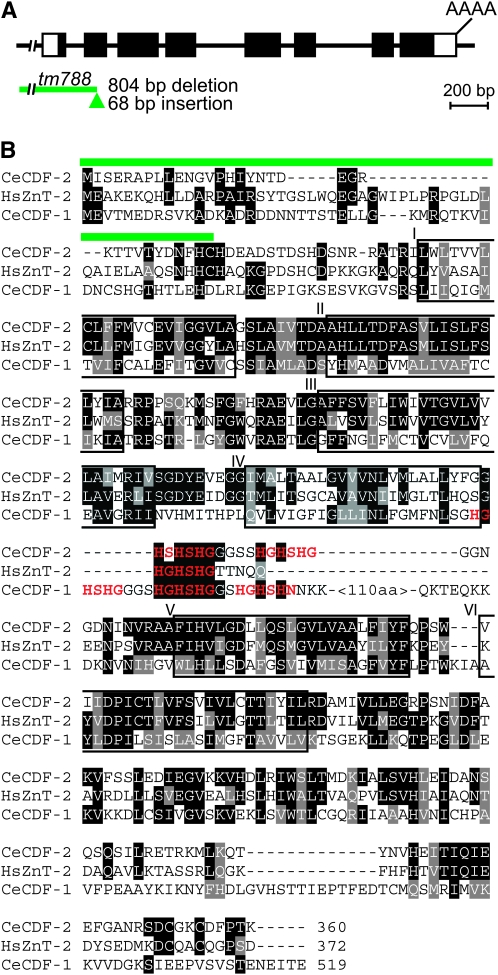
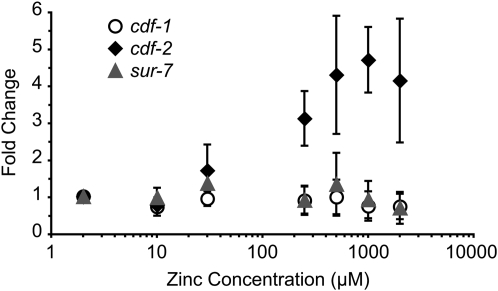
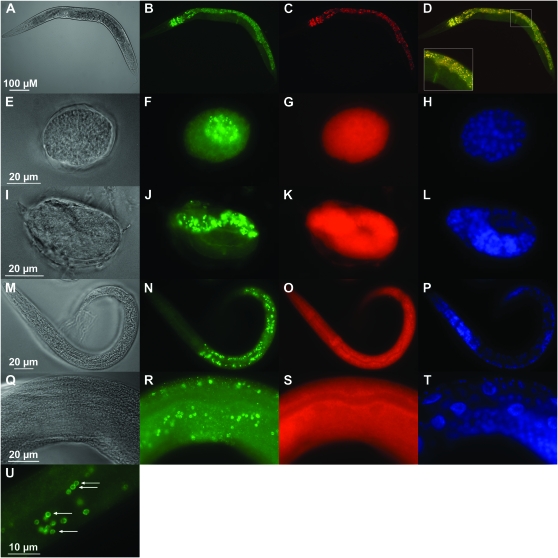
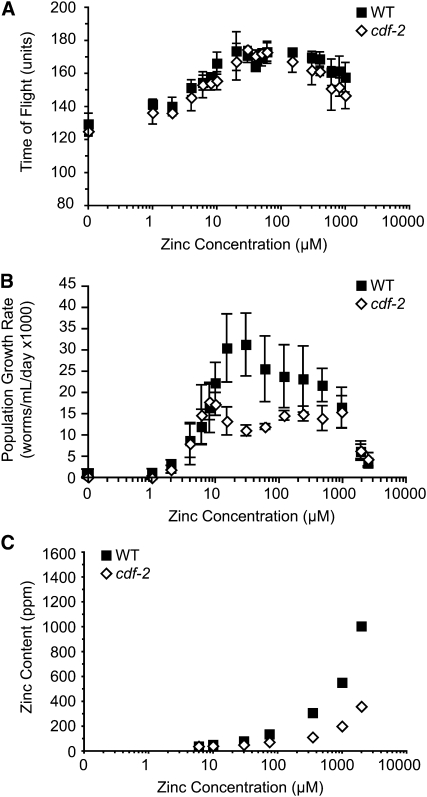
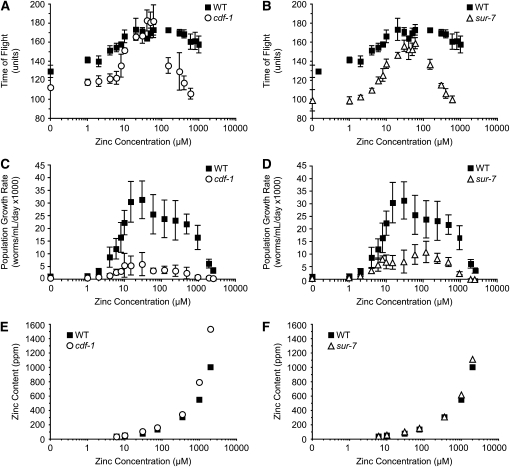
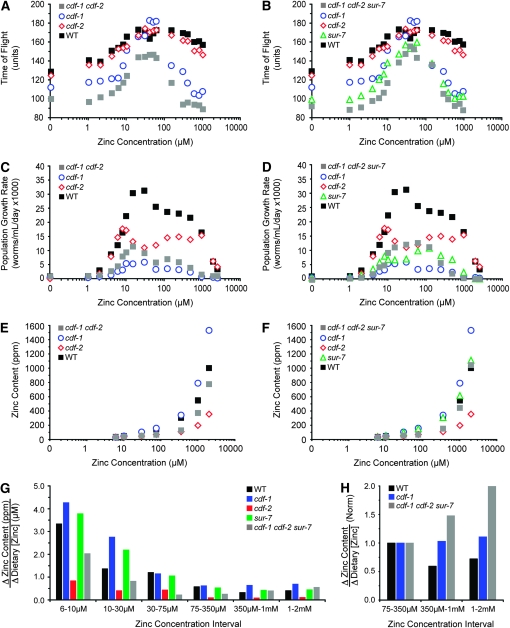
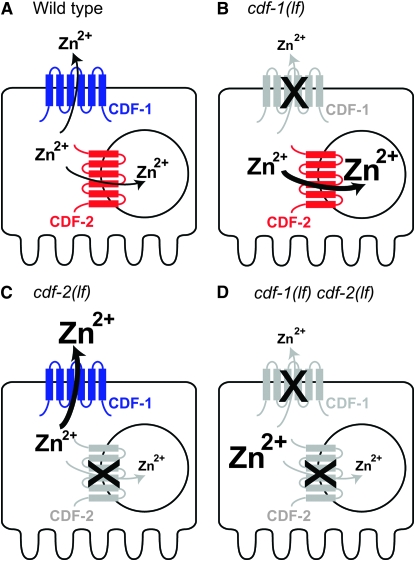
Zinc is essential for many cellular processes. To use Caenorhabditis elegans to study zinc metabolism, we developed culture conditions allowing full control of dietary zinc and methods to measure zinc content of animals. Dietary zinc dramatically affected growth and zinc content; wild-type worms survived from 7 microm to 1.3 mm dietary zinc, and zinc content varied 27-fold. We investigated cdf-2, which encodes a predicted zinc transporter in the cation diffusion facilitator family. cdf-2 mRNA levels were increased by high dietary zinc, suggesting cdf-2 promotes zinc homeostasis. CDF-2 protein was expressed in intestinal cells and localized to cytosolic vesicles. A cdf-2 loss-of-function mutant displayed impaired growth and reduced zinc content, indicating that CDF-2 stores zinc by transport into the lumen of vesicles. The relationships between three cdf genes, cdf-1, cdf-2, and sur-7, were analyzed in double and triple mutant animals. A cdf-1 mutant displayed increased zinc content, whereas a cdf-1 cdf-2 double mutant had intermediate zinc content, suggesting cdf-1 and cdf-2 have antagonistic functions. These studies advance C. elegans as a model of zinc metabolism and identify cdf-2 as a new gene that has a critical role in zinc storage.
Figures
References
-
- Andrews, G. K., H. Wang, S. K. Dey and R. D. Palmiter, 2004. Mouse zinc transporter 1 gene provides an essential function during early embryonic development. Genesis 40 74–81. - PubMed
-
- Beitel, G. J., S. G. Clark and H. R. Horvitz, 1990. Caenorhabditis elegans ras gene let-60 acts as a switch in the pathway of vulval induction. Nature 348 503–509. - PubMed
-
- Bruinsma, J. J., T. Jirakulaporn, A. J. Muslin and K. Kornfeld, 2002. Zinc ions and cation diffusion facilitator proteins regulate Ras-mediated signaling. Dev. Cell 2 567–578. - PubMed
-
- Bruinsma, J. J., D. L. Schneider, D. E. Davis and K. Kornfeld, 2008. Identification of mutations in Caenorhabditis elegans that cause resistance to high levels of dietary zinc and analysis using a genomewide map of single nucleotide polymorphisms scored by pyrosequencing. Genetics 179 811–828. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
