

doi: 10.2217/17460794.1.6.733.
HIV latency: present knowledge, future directions
Affiliations
- PMID: 19448843
- PMCID: PMC2682531
- DOI: 10.2217/17460794.1.6.733
Item in Clipboard
HIV latency: present knowledge, future directions
Future Virol.
2006.
Abstract
Current therapies do not eradicate HIV from infected patients. Indeed, HIV hides in a latent form insensitive to these therapies. Thus, one priority is to purge these latent reservoirs. But what mechanisms are responsible for latency and what are the reservoirs of latently infected cells? The present knowledge in terms of HIV latency is still incomplete and current therapeutic strategies fail to eradicate completely latently infected cells. What could the future bring?
Figures
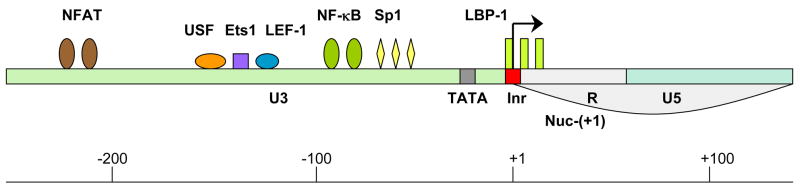
LTR is composed of three regions: 3′ untranslated region (U3) (green), transcription regulatory region (R) (grey) and 5′ untranslated region (U5) (blue). The following host TFs which bind to the HIV LTR are designated: NFAT (brown), USF (orange), Ets1 (purple), LEF-1 (blue), NF-kB (green), Sp1 (yellow) and LBP-1 (yellow-green). TATA box and initiator element are marked in grey and red, respectively. Nucleosome 1 location is from +1 to +155 (Nuc-(+1)). Arrow marks the transcription start site.
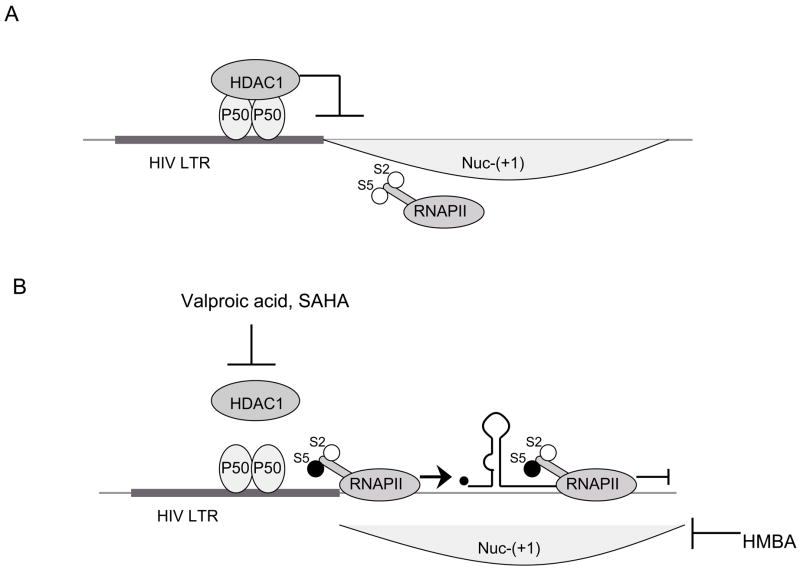
A) p50 homodimer bound to the LTR recruits HDAC1 which promotes deacetylation of histones. Thus, nucleosomes form compact chromatin structure and prevent transcription from the LTR. B) Inhibitors of HDAC1 (Valproic Acid and SAHA) which inhibit deacetylation of histones and HMBA relieve this repressive effect. Relaxation of chromatin structure allows the recruitment of preinitiation complex and phosphorylation of serine 5 of peptide repeats on CTD of RNAPII (S5). In this form, RNAPII successfully initiates transcription. However, transcriptionally active chromatin does not suffice for RNAPII to proceed to transcriptional elongation. For this step, RNAPII has to be phosphorylated on serine 2 of CTD peptide repeats (S2).
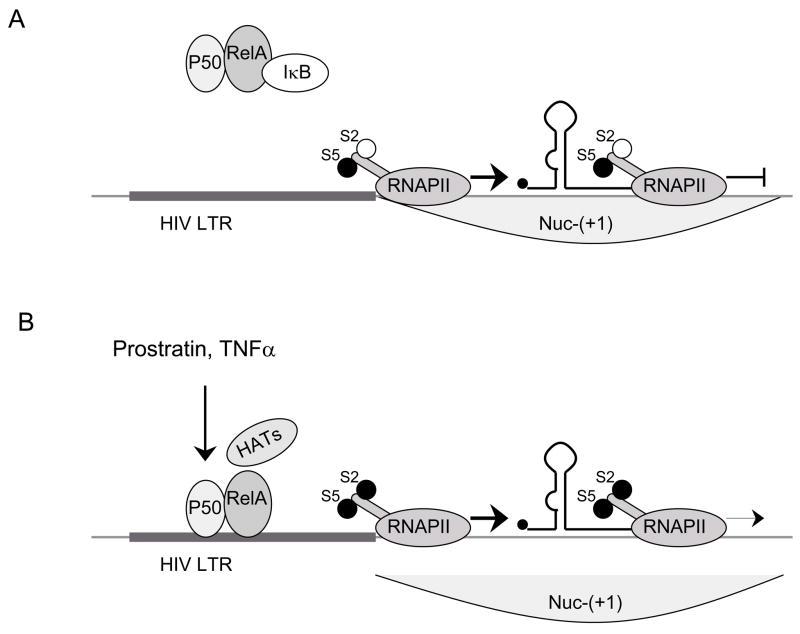
A) LTR is accessible for the recruitment of preinitiation complex, S5 is phosphorylated and RNAPII initiates transcription. Lack of positive TFs (non-active NF-κB is bound in the cytoplasm by IκB) results in insufficient transcriptional elongation. S2 is not phosphorylated and HATs are not recruited to the promoter (deacetylated histones in Nuc-(+1)). B) Prostratin and TNF-α increase levels of active NF-κB which results in the recruitment of HATs to the LTR. Possibly, P-TEFb is recruited to the promoter and it phosphorylates S2. Acetylation of histones by HATs relieve the transcriptional elongation block and leads to a suboptimal activation of transcriptional elongation.
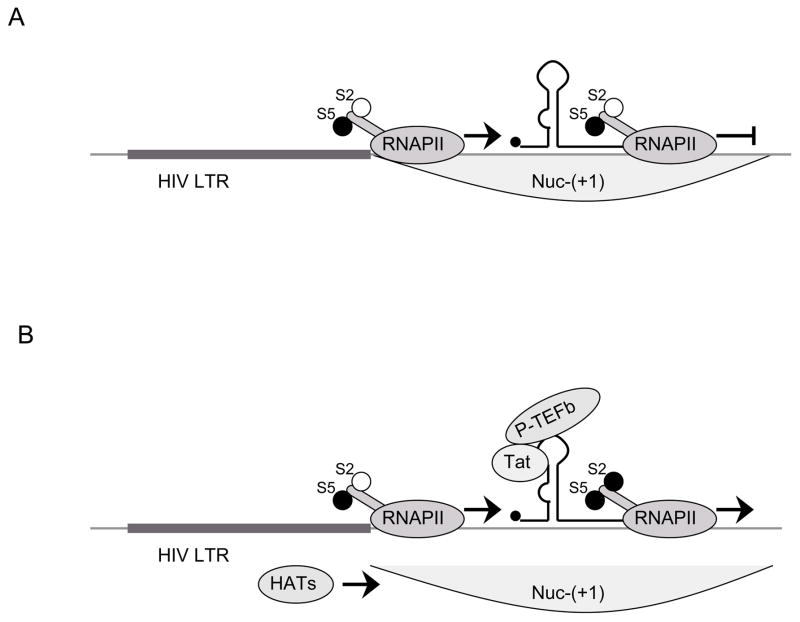
A) LTR is accessible for the recruitment of preinitiation complex, S5 is phosphorylated and RNAPII initiates transcription. Binding of NF-κB to the HIV LTR leads to the proceeding of RNAPII through HIV genome to some extent, however, lack of viral protein Tat prevents productive transcriptional elongation. B) Tat binds to TAR and recruits P-TEFb which in turn phosphorylates S2. Also, Tat recruits some of the HATs (e.g. PCAF) which acetylate histones following collapse of Nuc-(+1). This results in successful transcriptional elongation.
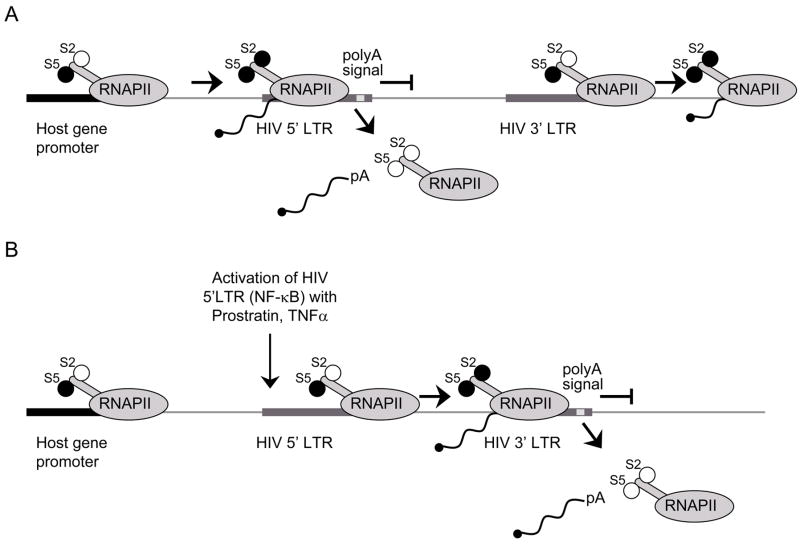
A) Active transcription from a host gene promoter interferes with transcription from downstream HIV 5′LTR. Polymerase ‘reads through’ the 5′LTR and dissociates from DNA at the polyadenylation signal in the 5′LTR resulting in a truncated form of mRNA. 3′ LTR is not occluded and is available for the recruitment of the preinitiation complex and proceeding of the transcription into the host gene. Transcription from the 3′LTR again results in a truncated form of mRNA. B) Activation of the 5′LTR (NF-κB, Tat) overcomes TI. Transcription is successfully initiated from 5′LTR. RNAPII proceeds through the HIV genome and terminates at the polyadenylation signal in 3′LTR. In this case, 3′LTR is occluded and there is no transcription from this LTR.
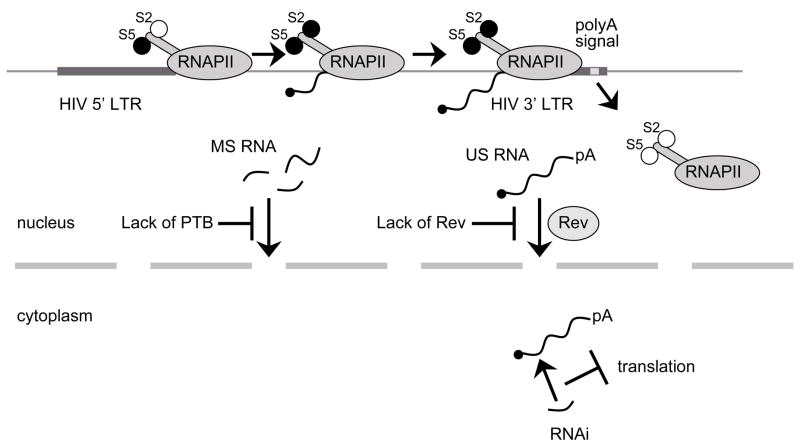
Viral genome is efficiently transcribed into full-length polyadenylated RNA. Aberrant localization of US and MS RNA: (i) without viral protein Rev US RNA are not protected from splicing and consequently, the full-length HIV RNA is not exported from the nucleus into the cytoplasm, (ii) too low levels of host protein PTB result in nuclear localization of MS RNA. Degradation of viral RNA: in the nucleus or in the cytoplasm viral RNA can be degraded by viral or host miRNA.
References
-
- Chun TW, Finzi D, Margolick J, Chadwick K, Schwartz D, Siliciano RF. In vivo fate of HIV-1-infected T cells: quantitative analysis of the transition to stable latency. Nat Med. 1995;1(12):1284–90. The first study in which stable form of latent HIV was found in purified resting CD4+ T-lymphocytes. - PubMed
-
- Finzi D, Blankson J, Siliciano JD, et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat Med. 1999;5(5):512–7. Demonstrates the importance of latent infection of resting CD4+ T cells for viral persistence and predicts the long time required for eradication of the latent reservoir. - PubMed
-
- Takahashi K, Wesselingh SL, Griffin DE, McArthur JC, Johnson RT, Glass JD. Localization of HIV-1 in human brain using polymerase chain reaction/in situ hybridization and immunocytochemistry. Ann Neurol. 1996;39(6):705–11. - PubMed
-
- Geijtenbeek TB, Torensma R, van Vliet SJ, et al. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell. 2000;100(5):575–85. - PubMed
-
- Igarashi T, Imamichi H, Brown CR, Hirsch VM, Martin MA. The emergence and characterization of macrophage-tropic SIV/HIV chimeric viruses (SHIVs) present in CD4+ T cell-depleted rhesus monkeys. J Leukoc Biol. 2003;74(5):772–80. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources