

An unusual recent expansion of the C-terminal domain of RNA polymerase II in primate malaria parasites features a motif otherwise found only in mammalian polymerases
- PMID: 19449052
- PMCID: PMC3622039
- DOI: 10.1007/s00239-009-9245-2
An unusual recent expansion of the C-terminal domain of RNA polymerase II in primate malaria parasites features a motif otherwise found only in mammalian polymerases
Abstract
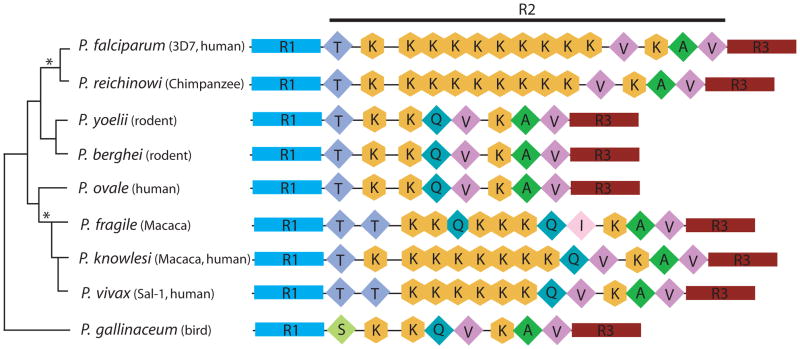
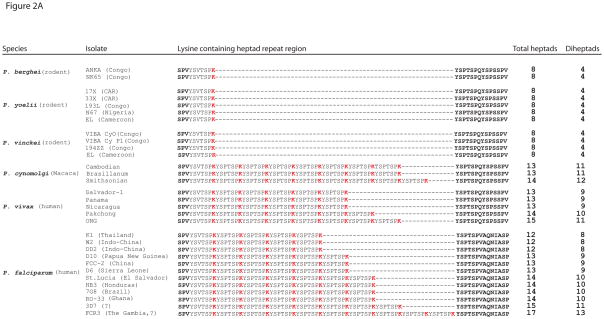
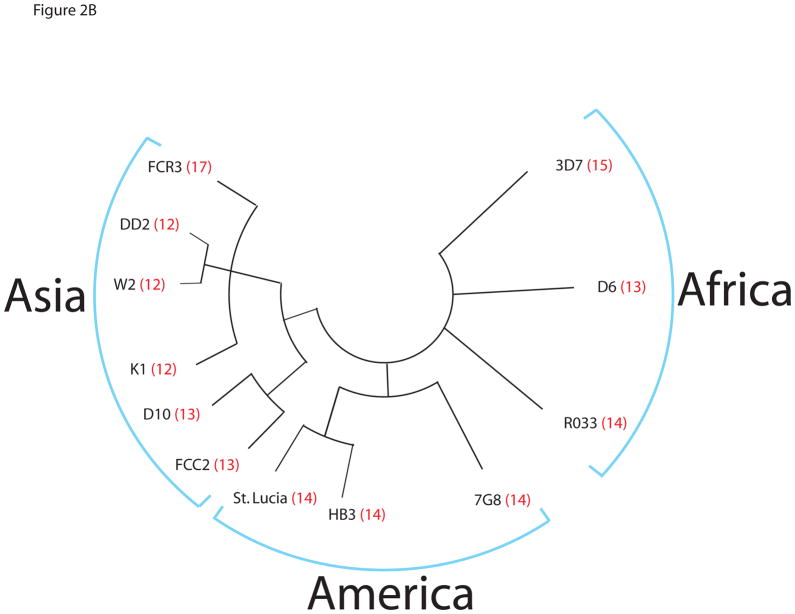
The tail of the enzyme RNA polymerase II is responsible for integrating the diverse events of gene expression in eukaryotes and is indispensable for life in yeast, fruit flies, and mice. The tail features a C-terminal domain (CTD), which is comprised of tandemly repeated Y(1)-S(2)-P(3)-T(4)-S(5)-P(6)-S(7) amino acid heptads that are highly conserved across evolutionary lineages, with all mammalian polymerases featuring 52 identical heptad repeats. However, the composition and function of protozoan CTDs remain less well understood. We find that malaria parasites (genus Plasmodium) display an unprecedented plasticity within the length and composition of their CTDs. The CTD in malaria parasites which infect human and nonhuman primates has expanded compared to closely related species that infect rodents or birds. In addition, this variability extends to different isolates within a single species, such as isolates of the human malaria parasite, Plasmodium falciparum. Our results indicate that expanded CTD heptads in malaria parasites correlates with parasitism of primates and provide the first demonstration of polymorphism of the RNA polymerase II CTD within a single species. The expanded set of CTD heptads feature lysine in the seventh position (Y(1)-S(2)-P(3)-T(4)-S(5)-P(6)-K(7)), a sequence only seen otherwise in the distal portion of mammalian polymerases. These observations raise new questions for the radiation of malaria parasites into diverse hosts and for the molecular evolution of RNA polymerase II.
Figures

Similar articles
-
RNA Polymerase II evolution and adaptations: Insights from Plasmodium and other parasitic protists.Infect Genet Evol. 2023 Nov;115:105505. doi: 10.1016/j.meegid.2023.105505. Epub 2023 Sep 23. Infect Genet Evol. 2023. PMID: 37748526
-
The C-terminal domain of RNA polymerase II of the malaria parasite Plasmodium berghei.Biochem Biophys Res Commun. 1991 Nov 14;180(3):1350-5. doi: 10.1016/s0006-291x(05)81344-0. Biochem Biophys Res Commun. 1991. PMID: 1840489
-
Evolution of lysine acetylation in the RNA polymerase II C-terminal domain.BMC Evol Biol. 2015 Mar 10;15:35. doi: 10.1186/s12862-015-0327-z. BMC Evol Biol. 2015. PMID: 25887984 Free PMC article.
-
Molecular evolution of the RNA polymerase II CTD.Trends Genet. 2008 Jun;24(6):289-96. doi: 10.1016/j.tig.2008.03.010. Epub 2008 May 9. Trends Genet. 2008. PMID: 18472177 Review.
-
An evolutionary perspective on the kinome of malaria parasites.Philos Trans R Soc Lond B Biol Sci. 2012 Sep 19;367(1602):2607-18. doi: 10.1098/rstb.2012.0014. Philos Trans R Soc Lond B Biol Sci. 2012. PMID: 22889911 Free PMC article. Review.
Cited by
-
Describing sequence-ensemble relationships for intrinsically disordered proteins.Biochem J. 2013 Jan 15;449(2):307-18. doi: 10.1042/BJ20121346. Biochem J. 2013. PMID: 23240611 Free PMC article. Review.
-
Erasing the epigenetic memory and beginning to switch--the onset of antigenic switching of var genes in Plasmodium falciparum.PLoS One. 2012;7(3):e34168. doi: 10.1371/journal.pone.0034168. Epub 2012 Mar 26. PLoS One. 2012. PMID: 22461905 Free PMC article.
-
Horizontal gene transfer of epigenetic machinery and evolution of parasitism in the malaria parasite Plasmodium falciparum and other apicomplexans.BMC Evol Biol. 2013 Feb 11;13:37. doi: 10.1186/1471-2148-13-37. BMC Evol Biol. 2013. PMID: 23398820 Free PMC article.
-
Vive la Différence: Exploiting the Differences between Rodent and Human Malarias.Trends Parasitol. 2020 Jun;36(6):504-511. doi: 10.1016/j.pt.2020.03.008. Epub 2020 Apr 16. Trends Parasitol. 2020. PMID: 32407681 Free PMC article. Review.
-
The serine/threonine phosphatases of apicomplexan parasites.Mol Microbiol. 2017 Oct;106(1):1-21. doi: 10.1111/mmi.13715. Epub 2017 Jun 14. Mol Microbiol. 2017. PMID: 28556455 Free PMC article. Review.
References
-
- Allison LA, Moyle M, Shales M, Ingles CJ. Extensive homology among the largest subunits of eukaryotic and prokaryotic RNA polymerases. Cell. 1985;42:599–610. - PubMed
-
- Aravind L, Iyer LM, Wellems TE, Miller LH. Plasmodium biology: Genomic gleanings. Cell. 2003;115:771–785. - PubMed
-
- Barron-Casella E, Corden JL. Conservation of the mammalian RNA polymerase II largest-subunit C-terminal domain. J Mol Evol. 1992;35:405–410. - PubMed
-
- Bienkiewicz EA, Moon WA, Woody RW. Conformation of the RNA polymerase II C-terminal domain: circular dichroism of long and short fragments. J Mol Biol. 2000;297:119–133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
