

Enzymatic methylation of DNA in cultured human cells studied by stable isotope incorporation and mass spectrometry
- PMID: 19449810
- PMCID: PMC3985286
- DOI: 10.1021/tx900027w
Enzymatic methylation of DNA in cultured human cells studied by stable isotope incorporation and mass spectrometry
Abstract
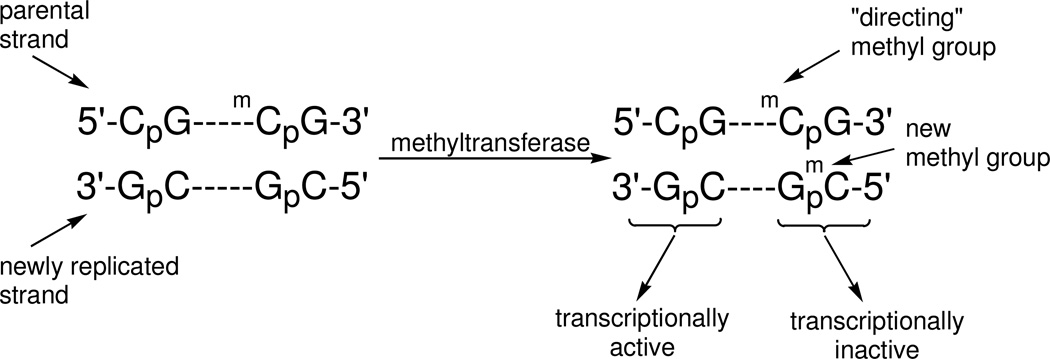
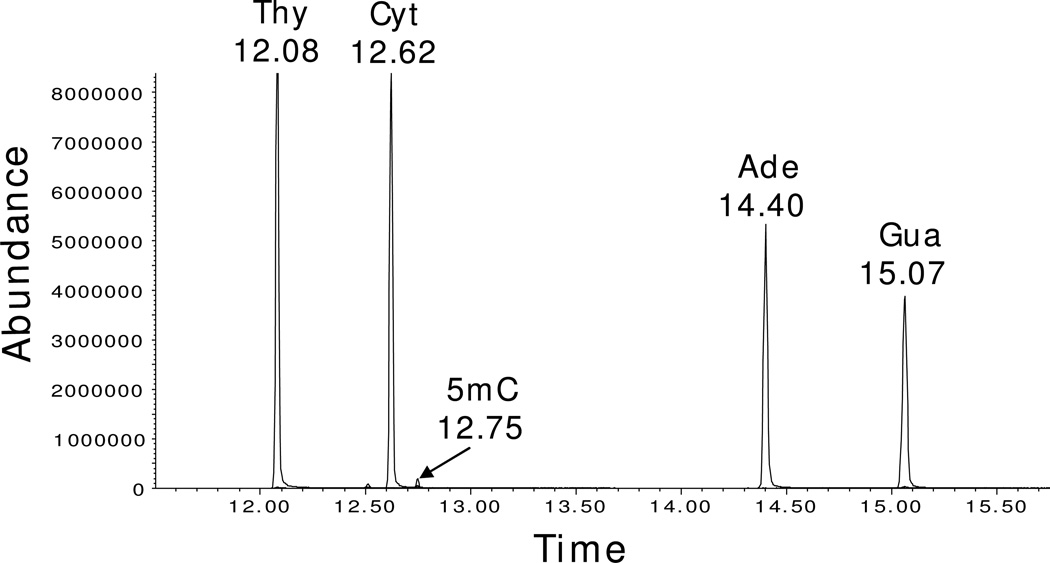
Enzymatic methylation of cytosine residues in DNA, in conjunction with covalent histone modifications, establishes an epigenetic code essential for the proper control of gene expression in higher organisms. Once established during cellular differentiation, the epigenetic code must be faithfully transmitted to progeny cells. However, epigenetic perturbations can be found in most if not all cancer cells, and the mechanisms leading to these changes are not well understood. In this paper, we describe a series of experiments aimed at understanding the dynamic process of DNA methylation that follows DNA replication. Cells in culture can be propagated in the presence of (15)N-enriched uridine, which labels the pyrimidine precursor pool as well as newly replicated DNA. Simultaneous culture in the presence of (2)H-enriched methionine results in labeling of newly methylated cytosine residues. An ensemble of 5-methylcytosine residues differing in the degree of isotopic enrichment is generated, which can be examined by mass spectrometry. Using this method, we demonstrate that the kinetics of both DNA replication and methylation of newly replicated DNA are indistinguishable. The majority of methylation following DNA replication is shown to occur on the newly synthesized DNA. The method reported here does, however, suggest an unexpected methylation of parental DNA during DNA replication, which might indicate a previously undescribed chromatin remodeling process. The method presented here will be useful in monitoring the dynamic process of DNA methylation and will allow a more detailed understanding of the mechanisms of clinically used methylation inhibitors and environmental toxicants.
Figures







Similar articles
-
Characteristics of enzymatic DNA methylation in cultured cells of human and hamster origin, and the effect of DNA replication inhibition.Biochim Biophys Acta. 1982 Jan 26;696(1):15-22. doi: 10.1016/0167-4781(82)90004-5. Biochim Biophys Acta. 1982. PMID: 7200803
-
Widespread existence of cytosine methylation in yeast DNA measured by gas chromatography/mass spectrometry.Anal Chem. 2012 Aug 21;84(16):7249-55. doi: 10.1021/ac301727c. Epub 2012 Aug 1. Anal Chem. 2012. PMID: 22852529 Free PMC article.
-
Examination of the DNA substrate selectivity of DNA cytosine methyltransferases using mass tagging.Nucleic Acids Res. 2000 Sep 15;28(18):3594-9. doi: 10.1093/nar/28.18.3594. Nucleic Acids Res. 2000. PMID: 10982881 Free PMC article.
-
5-methylcytosine and its derivatives.Adv Clin Chem. 2014;67:151-87. doi: 10.1016/bs.acc.2014.09.003. Epub 2014 Nov 4. Adv Clin Chem. 2014. PMID: 25735861 Review.
-
Detecting and interpreting DNA methylation marks.Curr Opin Struct Biol. 2018 Dec;53:88-99. doi: 10.1016/j.sbi.2018.06.004. Epub 2018 Jul 19. Curr Opin Struct Biol. 2018. PMID: 30031306 Free PMC article. Review.
Cited by
-
Polymerase incorporation and miscoding properties of 5-chlorouracil.Chem Res Toxicol. 2010 Apr 19;23(4):740-8. doi: 10.1021/tx900302j. Chem Res Toxicol. 2010. PMID: 20104909 Free PMC article.
-
Untargeted 2D NMR Metabolomics of [13C-methyl]Methionine-Labeled Tumor Models Reveals the Non-DNA Methylome and Provides Clues to Methyl Metabolism Shift during Tumor Progression.J Proteome Res. 2022 Apr 1;21(4):940-952. doi: 10.1021/acs.jproteome.1c00778. Epub 2022 Feb 23. J Proteome Res. 2022. PMID: 35196455 Free PMC article.
-
Analysis of glucose-derived amino acids involved in one-carbon and cancer metabolism by stable-isotope tracing gas chromatography mass spectrometry.Anal Biochem. 2019 Feb 1;566:1-9. doi: 10.1016/j.ab.2018.10.026. Epub 2018 Oct 26. Anal Biochem. 2019. PMID: 30409761 Free PMC article.
-
Glioblastoma and Methionine Addiction.Int J Mol Sci. 2022 Jun 28;23(13):7156. doi: 10.3390/ijms23137156. Int J Mol Sci. 2022. PMID: 35806160 Free PMC article. Review.
-
Mass spectrometric studies on epigenetic interaction networks in cell differentiation.J Biol Chem. 2011 Apr 15;286(15):13657-68. doi: 10.1074/jbc.M110.204800. Epub 2011 Feb 18. J Biol Chem. 2011. PMID: 21335548 Free PMC article.
References
-
- Razin A, Riggs AD. DNA methylation and gene function. Science. 1980;210:604–610. - PubMed
-
- Jones PA, Taylor SM. Cellular differentiation, cytidine analogs and DNA methylation. Cell. 1980;20:85–93. - PubMed
-
- Singal R, Ginder GD. DNA methylation. Blood. 1999;93:4059–4070. - PubMed
-
- Ehrlich M, Wang RY-H. 5-Methylcytosine in eukaryotic DNA. Science. 1981;212:1350–1357. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
