

Hematopoietic stem cell development is dependent on blood flow
- PMID: 19450519
- PMCID: PMC2722870
- DOI: 10.1016/j.cell.2009.04.023
Hematopoietic stem cell development is dependent on blood flow
Abstract
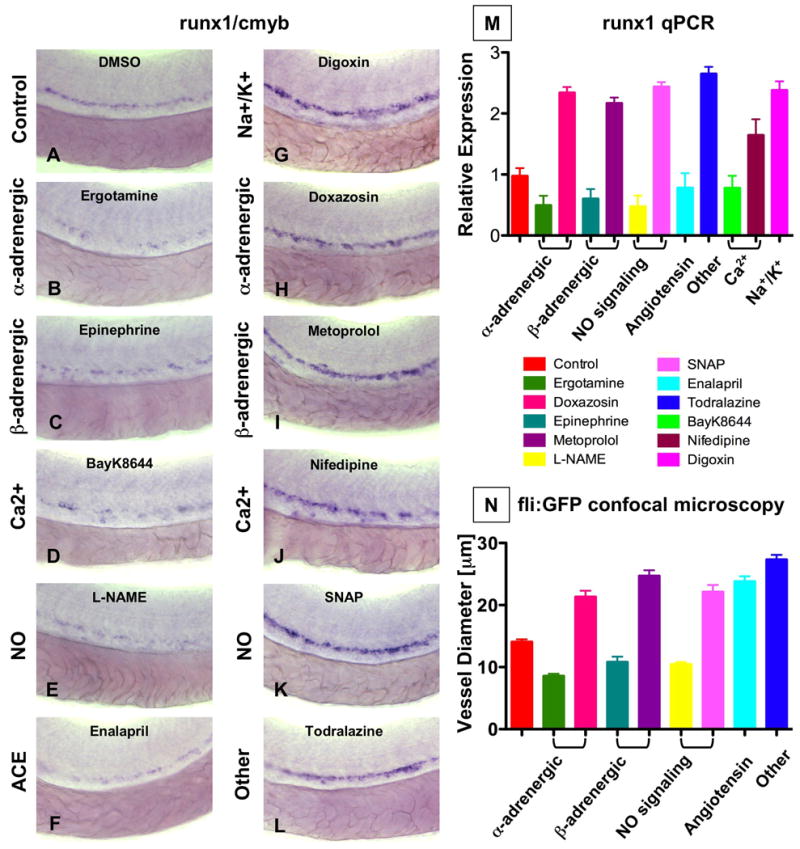
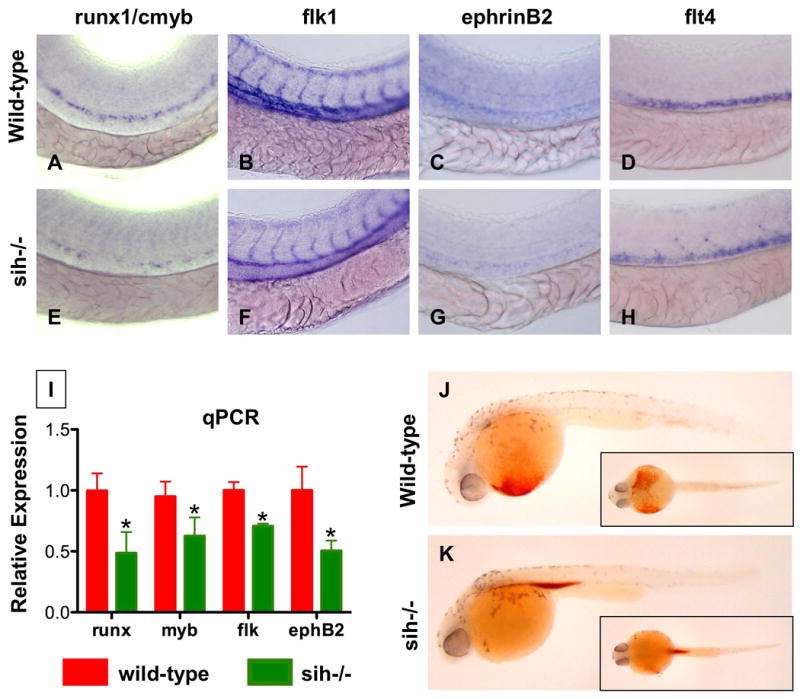
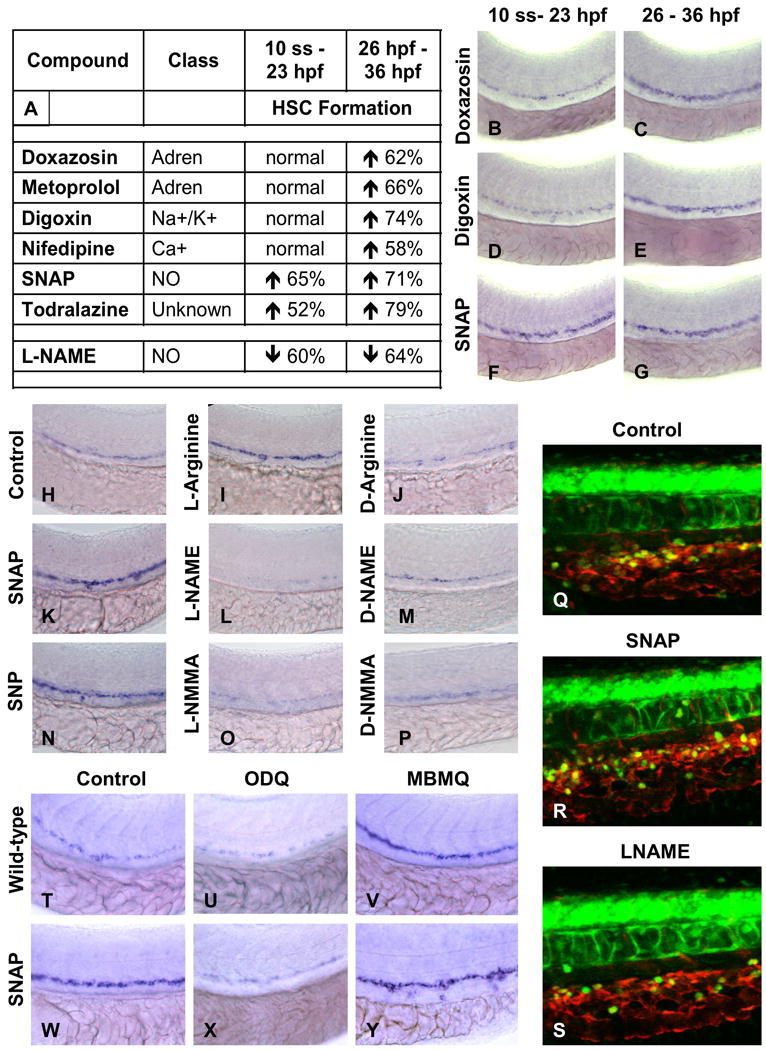
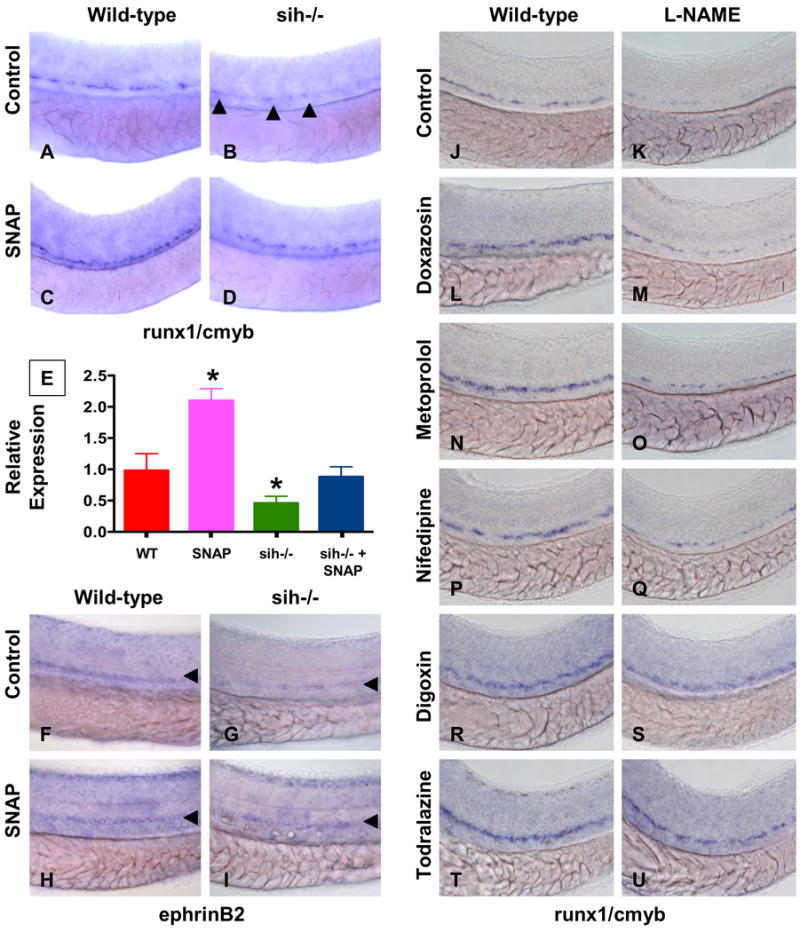
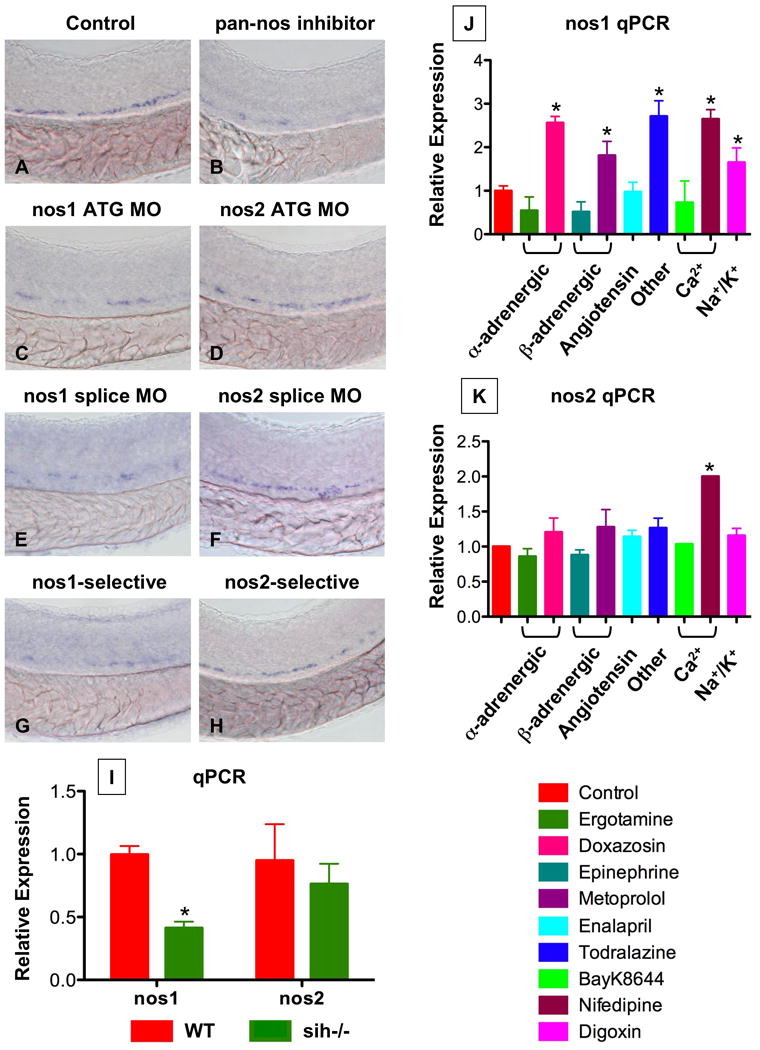
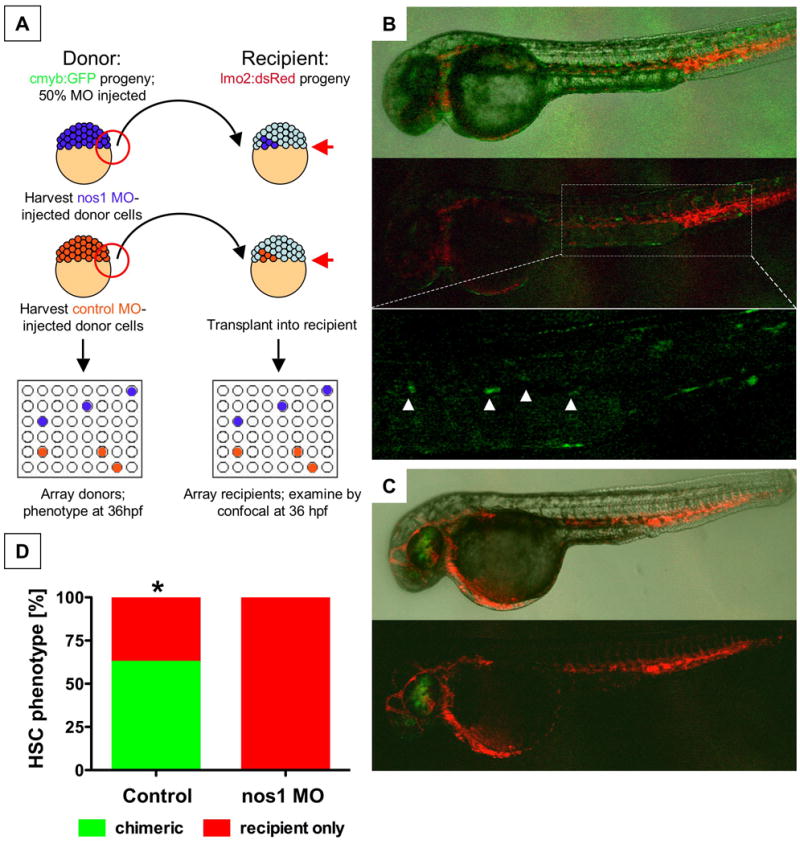
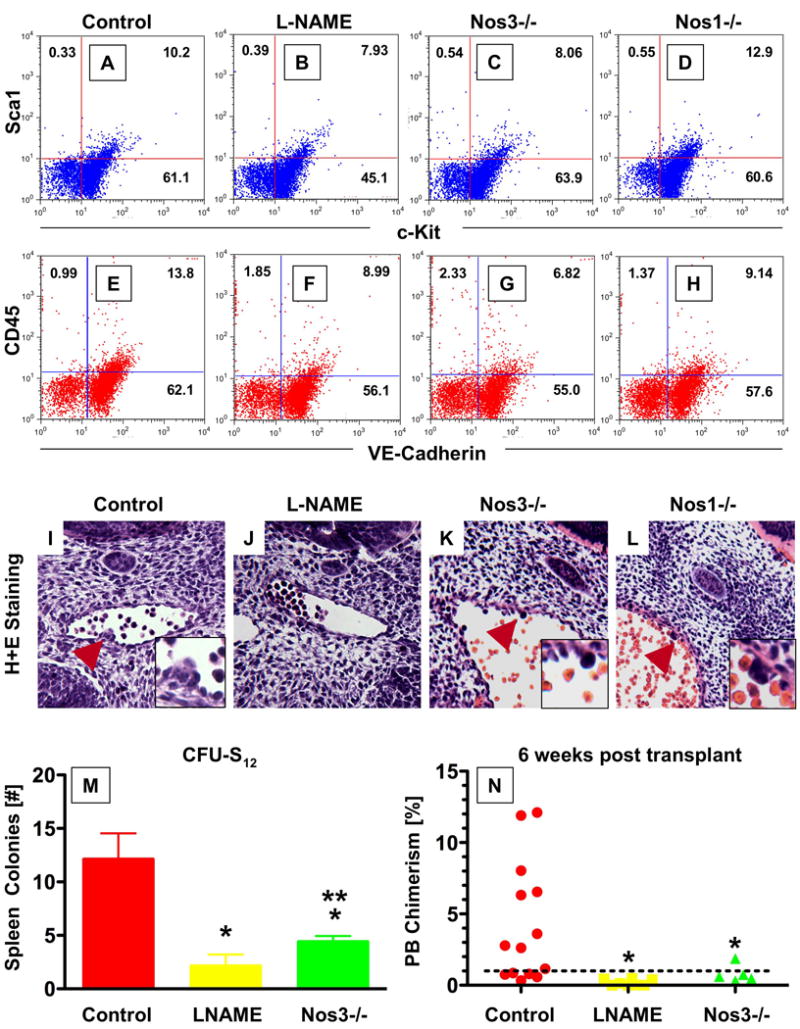
During vertebrate embryogenesis, hematopoietic stem cells (HSCs) arise in the aorta-gonads-mesonephros (AGM) region. We report here that blood flow is a conserved regulator of HSC formation. In zebrafish, chemical blood flow modulators regulated HSC development, and silent heart (sih) embryos, lacking a heartbeat and blood circulation, exhibited severely reduced HSCs. Flow-modifying compounds primarily affected HSC induction after the onset of heartbeat; however, nitric oxide (NO) donors regulated HSC number even when treatment occurred before the initiation of circulation, and rescued HSCs in sih mutants. Morpholino knockdown of nos1 (nnos/enos) blocked HSC development, and its requirement was shown to be cell autonomous. In the mouse, Nos3 (eNos) was expressed in HSCs in the AGM. Intrauterine Nos inhibition or embryonic Nos3 deficiency resulted in a reduction of hematopoietic clusters and transplantable murine HSCs. This work links blood flow to AGM hematopoiesis and identifies NO as a conserved downstream regulator of HSC development.
Figures
References
-
- Aicher A, Heeschen C, Mildner-Rihm C, Urbich C, Ihling C, Technau-Ihling K, Zeiher AM, Dimmeler S. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat Med. 2003;9:1370–1376. - PubMed
-
- Burggren WW. What is the purpose of the embryonic heart beat? Or how facts can ultimately prevail over physiological dogma. Physiol Biochem Zool. 2004;77:333–345. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
