

TFIIH kinase places bivalent marks on the carboxy-terminal domain of RNA polymerase II
- PMID: 19450536
- PMCID: PMC2757088
- DOI: 10.1016/j.molcel.2009.04.016
TFIIH kinase places bivalent marks on the carboxy-terminal domain of RNA polymerase II
Abstract
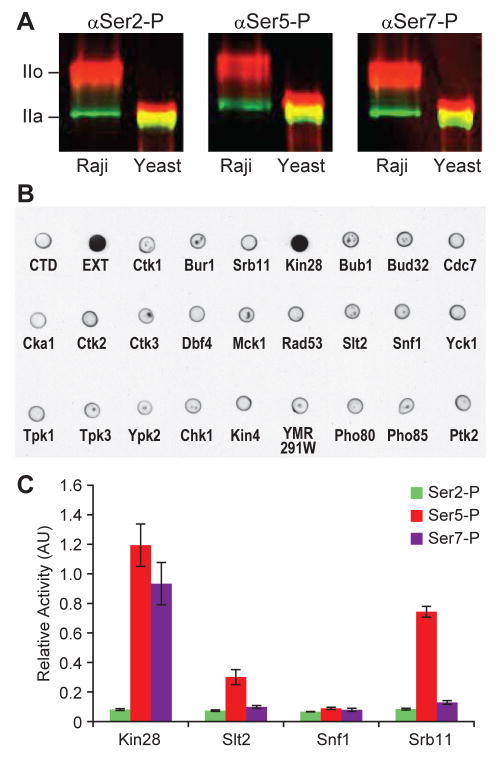
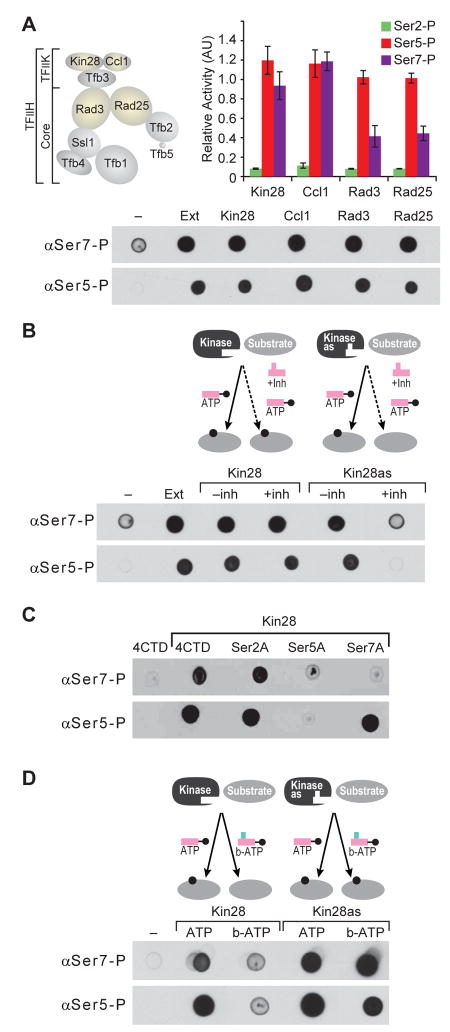
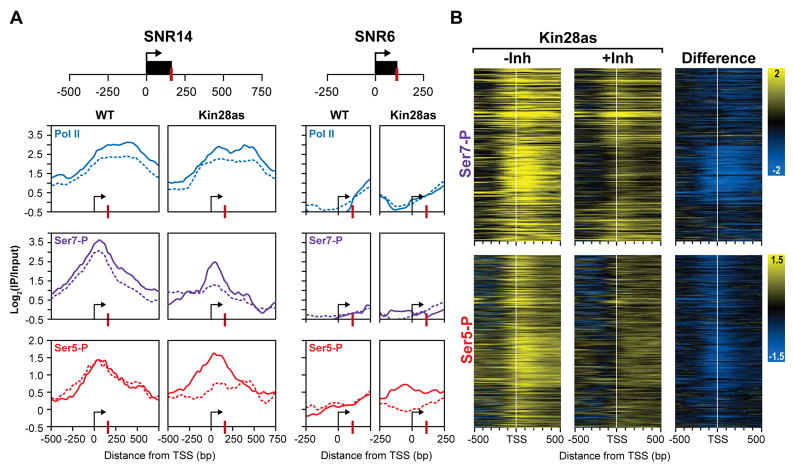
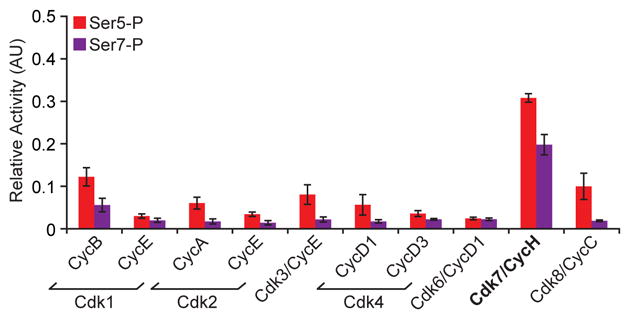
Posttranslational modifications of the carboxy-terminal domain (CTD) of the largest subunit of RNA polymerase II (Pol II) specify a molecular recognition code that is deciphered by proteins involved in RNA biogenesis. The CTD is comprised of a repeating heptapeptide (Y(1)S(2)P(3)T(4)S(5)P(6)S(7)). Recently, phosphorylation of serine 7 was shown to be important for cotranscriptional processing of two snRNAs in mammalian cells. Here we report that Kin28/Cdk7, a subunit of the evolutionarily conserved TFIIH complex, is a Ser7 kinase. The ability of Kin28/Cdk7 to phosphorylate Ser7 is particularly surprising because this kinase functions at promoters of protein-coding genes, rather than being restricted to promoter-distal regions of snRNA genes. Kin28/Cdk7 is also known to phosphorylate Ser5 residues of the CTD at gene promoters. Taken together, our results implicate the TFIIH kinase in placing bivalent Ser5 and Ser7 marks early in gene transcription. These bivalent CTD marks, in concert with cues within nascent transcripts, specify the cotranscriptional engagement of the relevant RNA processing machinery.
Figures
References
-
- Ahn SH, Kim M, Buratowski S. Phosphorylation of serine 2 within the RNA Polymerase II C-terminal domain couples transcription and 3′ end processing. Mol Cell. 2004;13:67–76. - PubMed
-
- Baillat D, Hakimi MA, Näär AM, Shilatifard A, Cooch N, Shiekhattar R. Integrator, a multiprotein mediator of small nuclear RNA processing, associates with the C-terminal repeat of RNA Polymerase II. Cell. 2005;123:265–276. - PubMed
-
- Bishop AC, Ubersax JA, Petsch DT, Matheos DP, Gray NS, Blethrow J, Shimizu E, Tsien JZ, Schultz PG, Rose MD, et al. A chemical switch for inhibitor-sensitive alleles of any protein kinase. Nature. 2000;407:395–401. - PubMed
-
- Bishop AC, Buzko O, Shokat KM. Magic bullets for protein kinases. Trends Cell Biol. 2001;11:167–172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
