

Androgens enhance plasticity of an electric communication signal in female knifefish, Brachyhypopomus pinnicaudatus
- PMID: 19450600
- PMCID: PMC2722804
- DOI: 10.1016/j.yhbeh.2009.05.005
Androgens enhance plasticity of an electric communication signal in female knifefish, Brachyhypopomus pinnicaudatus
Abstract
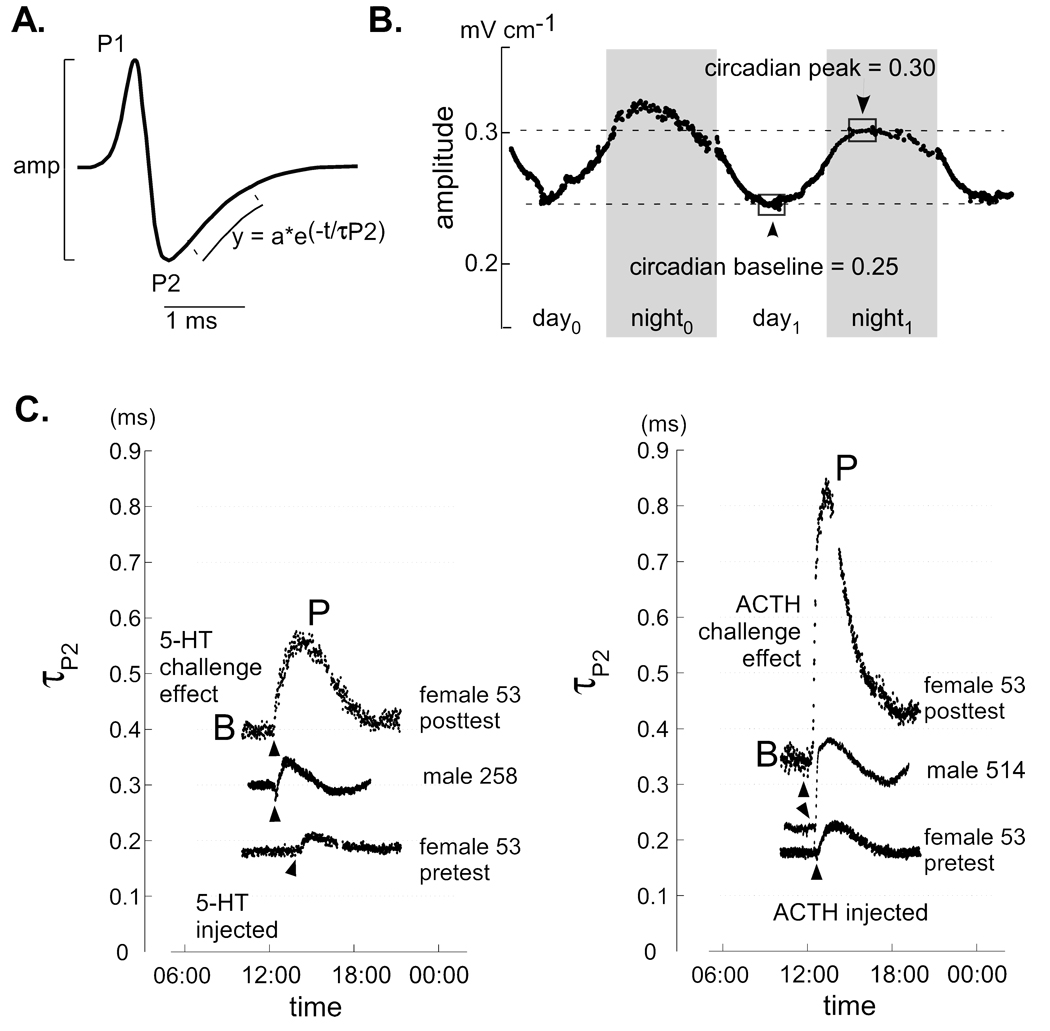
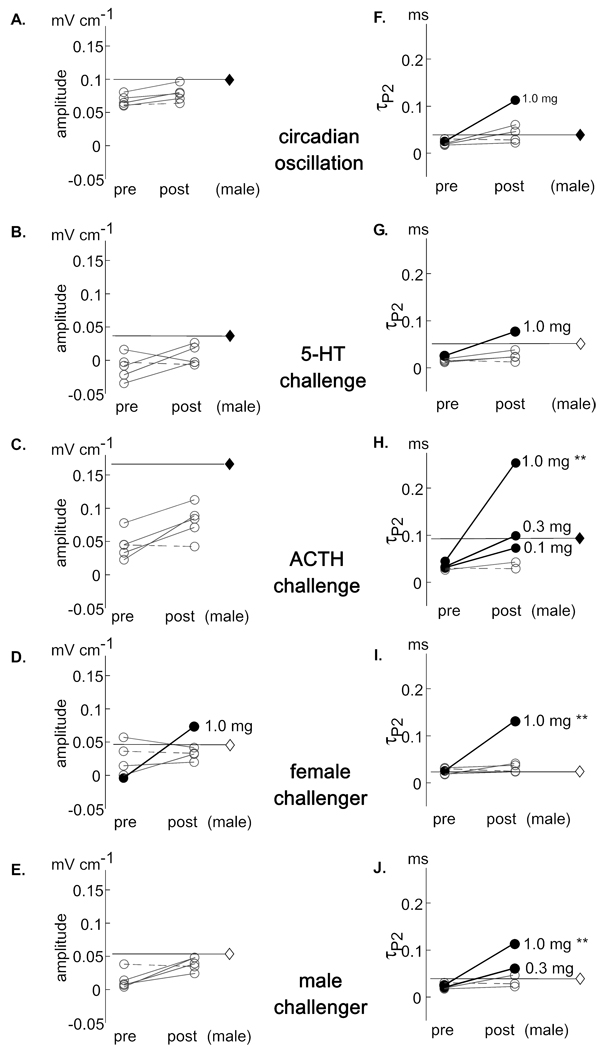
Sex steroids were initially defined by their actions shaping sexually dimorphic behavioral patterns. More recently scientists have begun exploring the role of steroids in determining sex differences in behavioral plasticity. We investigated the role of androgens in potentiating circadian, pharmacological, and socially-induced plasticity in the amplitude and duration of electric organ discharges (EODs) of female gymnotiform fish. We first challenged female fish with injections of serotonin (5-HT) and adrenocorticotropic hormone (ACTH), and with social encounters with female and male conspecifics to characterize females' pre-implant responses to each treatment. Each individual was then implanted with a pellet containing dihydrotestosterone (DHT) concentrations of 0.0, 0.03, 0.1, 0.3, or 1.0 mg 10 g(-1) body weight. We then repeated all challenges and compared each female's pre- and post-implant responses. The highest implant dose enhanced EOD duration modulations in response to all challenge types, responses to male challenge were also greater at the second highest dose, and responses to ACTH challenge were enhanced in females receiving all but the smallest dose (and blank) implants. Alternatively, amplitude modulations were enhanced only during female challenges and only when females received the highest DHT dose. Our results highlight the differential regulation of EOD duration and amplitude, and suggest that DHT enhanced the intrinsic plasticity of the electrogenic cells that produce the EOD rather than modifying behavioral phenotypes. The relative failure of DHT to enhance EOD amplitude plasticity also implies that factors other than androgens are involved in regulating/promoting male-typical EOD circadian rhythms and waveform modulations displayed in social contexts.
Figures
References
-
- Adkins-Regan E. Adler N. Neuroendocrinology of Reproduction. New York: Plenum Press; 1981. Early organizational effects of hormones: An evolutionary perspective; pp. 159–221.
-
- Adkins-Regan E. Hormonal mechanisms of mate choice. Am. Zool. 1998;38:166–178.
-
- Andersson MB. Princeton, N.J: Princeton University Press; 1994. Sexual selection.
-
- Arnold AP, Breedlove SM. Organizational and activational effects of sex steroids on brain and behavior: a reanalysis. Horm. Behav. 1985;19:469–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
