

Food-web structure and ecosystem services: insights from the Serengeti
- PMID: 19451118
- PMCID: PMC2685430
- DOI: 10.1098/rstb.2008.0287
Food-web structure and ecosystem services: insights from the Serengeti
Abstract
The central organizing theme of this paper is to discuss the dynamics of the Serengeti grassland ecosystem from the perspective of recent developments in food-web theory. The seasonal rainfall patterns that characterize the East African climate create an annually oscillating, large-scale, spatial mosaic of feeding opportunities for the larger ungulates in the Serengeti; this in turn creates a significant annual variation in the food available for their predators. At a smaller spatial scale, periodic fires during the dry season create patches of highly nutritious grazing that are eaten in preference to the surrounding older patches of less palatable vegetation. The species interactions between herbivores and plants, and carnivores and herbivores, are hierarchically nested in the Serengeti food web, with the largest bodied consumers on each trophic level having the broadest diets that include species from a large variety of different habitats in the ecosystem. The different major habitats of the Serengeti are also used in a nested fashion; the highly nutritious forage of the short grass plains is available only to the larger migratory species for a few months each year. The longer grass areas, the woodlands and kopjes (large partially wooded rocky islands in the surrounding mosaic of grassland) contain species that are resident throughout the year; these species often have smaller body size and more specialized diets than the migratory species. Only the larger herbivores and carnivores obtain their nutrition from all the different major habitat types in the ecosystem. The net effect of this is to create a nested hierarchy of subchains of energy flow within the larger Serengeti food web; these flows are seasonally forced by rainfall and operate at different rates in different major branches of the web. The nested structure that couples sequential trophic levels together interacts with annual seasonal variation in the fast and slow chains of nutrient flow in a way that is likely to be central to the stability of the whole web. If the Serengeti is to be successfully conserved as a fully functioning ecosystem, then it is essential that the full diversity of natural habitats be maintained within the greater Serengeti ecosystem. The best way to do this is by controlling the external forces that threaten the boundaries of the ecosystem and by balancing the economic services the park provides between local, national and international needs. I conclude by discussing how the ecosystem services provided by the Serengeti are driven by species on different trophic levels. Tourism provides the largest financial revenue to the national economy, but it could be better organized to provide more sustained revenue to the park. Ultimately, ecotourism needs to be developed in ways that take lessons from the structure of the Serengeti food webs, and in ways that provide tangible benefits to people living around the park while also improving the experience of all visitors.
Figures
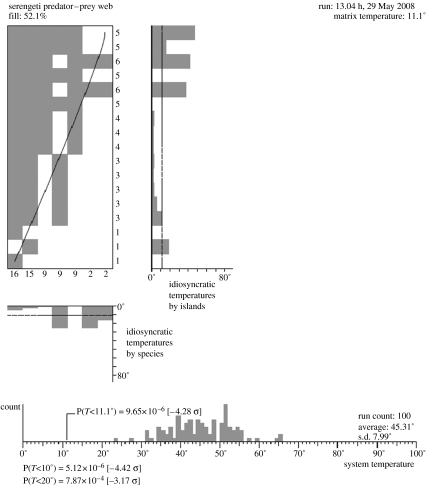
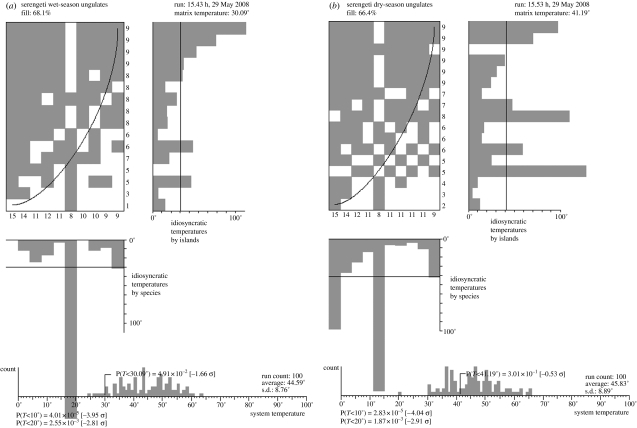
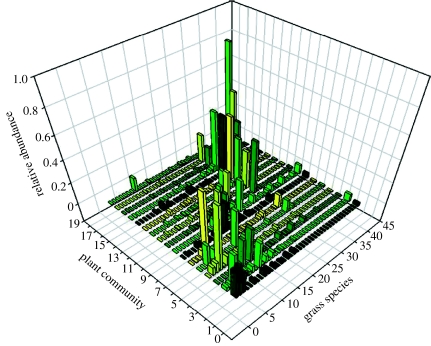
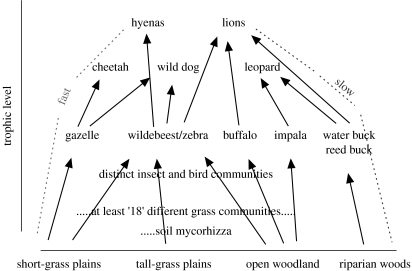
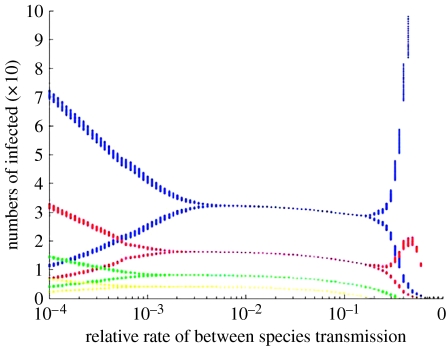
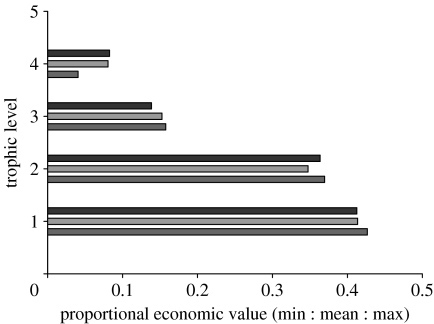
References
-
- Allesina S., Pascual M. Network structure, predator–prey modules, and stability in large food webs. Theor. Ecol. 2008;1:55–64. doi:10.1007/s12080-007-0007-8 - DOI
-
- Anderson R.M. Parasite pathogenicity and the depression of host population equilibria. Nature. 1979;279:150–152. doi:10.1038/279150a0 - DOI
-
- Anderson G.D., Talbot L.M. Soil factors affecting the distribution of the grassland types and their utilization by wild animals on the Serengeti plains, Tanganyika. J. Ecol. 1965;53:33–56. doi:10.2307/2257564 - DOI
-
- Anderson T.M., Ritchie M.E., Mayemba E., Eby S., Grace J.B., McNaughton S.J. Forage nutritive quality in the Serengeti ecosystem: the roles of fire and herbivory. Am. Nat. 2007;170:343–357. doi:10.1086/520120 - DOI - PubMed
-
- Anderson T.M., Dempewolf J., Metzger K.L., Reed D.N., Serneels S. Generation and maintenance of heterogeneity in the Serengeti ecosystem. In: Sinclair A.R.E., Packer C., Mduma S.A.R., Fryxell J.M., editors. Serengeti III. Huamn impacts on ecosystem dynamics. University of Chicago Press; Chicago, IL: 2008. pp. 135–182.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources