

doi: 10.1101/gad.510809.
The H3K27me3 demethylase JMJD3 contributes to the activation of the INK4A-ARF locus in response to oncogene- and stress-induced senescence
Affiliations
- PMID: 19451217
- PMCID: PMC2685535
- DOI: 10.1101/gad.510809
Item in Clipboard
The H3K27me3 demethylase JMJD3 contributes to the activation of the INK4A-ARF locus in response to oncogene- and stress-induced senescence
Genes Dev.
.
Abstract
The tumor suppressor proteins p16INK4A and p14ARF, encoded by the INK4A-ARF locus, are key regulators of cellular senescence. The locus is epigenetically silenced by the repressive H3K27me3 mark in normally growing cells, but becomes activated in response to oncogenic stress. Here, we show that expression of the histone H3 Lys 27 (H3K27) demethylase JMJD3 is induced upon activation of the RAS-RAF signaling pathway. JMJD3 is recruited to the INK4A-ARF locus and contributes to the transcriptional activation of p16INK4A in human diploid fibroblasts. Additionally, inhibition of Jmjd3 expression in mouse embryonic fibroblasts results in suppression of p16Ink4a and p19Arf expression and in their immortalization.
Figures

JMJD3 expression is induced by oncogenic BRAF and HRASV12. (A–C) Expression levels of p16INK4A and p14ARF (A), JMJD3 and UTX (B), and EZH2 and SUZ12 (C) in TIG3 BRAF-ER at different times following OHT treatment determined by RT-qPCR. Data presented in A–C are derived from the same experiment, but are representative of at least three independent experiments. (D) Expression levels of INK4A, ARF, and JMJD3 in BJ BRAF-ER at different times (0, 2, 3, 4 d) following OHT treatment, determined by RT-qPCR. (E) Protein levels of JMJD3, p16INK4A, and EZH2 determined by Western blotting following the activation of BRAF. (F,G) Expression levels of INK4A, ARF, and JMJD3 in TIG3-hTERT and BJ-hTERT cells transduced with HRASV12-encoding retrovirus. After selection, expression levels were determined by RT-qPCR. (H) Protein levels of JMJD3 in TIG3-hTERT cells at different time points after treatment with 50 J/m2 UV irradiation.

Induction of p16INK4A expression in response to BRAF is regulated by JMJD3. (A) Reporter assays using lysates from BRAF-ER U2OS cells transfected with various human JMJD3 promoter/luciferase reporter gene constructs with or without OHT stimulation. OHT or mock was added 36 h after transfection, and cells were harvested after 48 h and assayed. (B) Schematic representation of the human JMJD3 locus. Gray boxes indicate the size and position of the different promoter constructs (P1, P2, P3, etc.) used. Arrows indicate the ESC-TSS, and MF-TSS. (C–F) TIG3 BRAF-ER cells were transduced with shRNA plasmids targeting JMJD3 [shJMJD3(3302)] or control shRNA. Following selection, expression levels of JMJD3, INK4A, UTX, and EZH2 mRNA were monitored by RT-qPCR after OHT treatment. (G) The protein levels of JMJD3, p16INK4A, UTX, and EZH2 determined by Western blotting following OHT treatment. Data presented in C–F are derived from the same experiment and are representative of at least three independent experiments. Error bars on qPCR experiments and reporter assays indicate standard deviations.

Overexpression of JMJD3 results in increased expression of INK4A in MEFs and TIG3 cells. (A) Wild-type, Tp53−/−, Arf−/−, and Ink4a–Arf−/− cells were infected with pBabe constructs expressing a HA-tagged version of the catalytically active fragment of JMJD3 (JMJD3s), as well as the catalytically inactive version (JMJD3sMT, or D3sMT in figure). Mock refers to cells infected with empty pBabe vector. The expression levels of Ink4a and Arf were measured by RT-qPCR. Data are plotted as relative to mock-infected cells. (B) The expression level of INK4A was measured by RT-qPCR in TIG3-hTERT cells infected with mock, JMJD3s, and JMJD3sMT. Data are plotted as relative to mock-infected cells. (C) Growth curves using MEFs with the indicated genotypes infected with mock, JMJD3s, and JMJD3sMT. Y-axes are log2 scale.
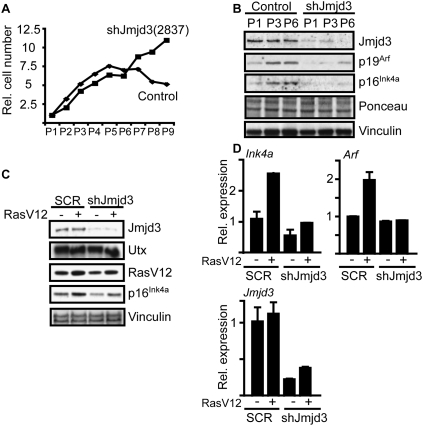
shRNA-mediated depletion of Jmjd3 inhibits stress-induced expression of p16Ink4a and p19Arf and leads to immortalization of primary MEFs. Primary MEFs were transduced with shRNA plasmids targeting Jmjd3 [shJmjd3(2837)] or control shRNA. (A) Jmjd3 depletion leads to immortalization of MEFs. The MEFs were passaged every 3 d and the relative cell number was plotted against the passage number. (B) Western blotting of cell lysates prepared at the indicated passages using antibodies specific for the indicated proteins. (C) MEFs were coinfected with a lentiviral RasV12 expression construct and shJmjd3(2837) or control shRNA. (D) Cells were grown for 1–2 wk and expression levels of p16Ink4a, p19Arf, Jmjd3, and RasV12 were measured by immunoblotting and by RT-qPCR.
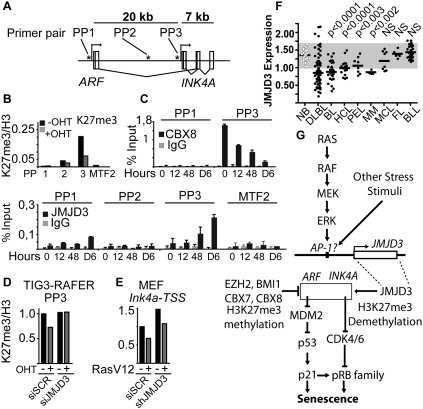
JMJD3 associates with the INK4A–ARF locus. (A) Schematic presentation of the INK4A–ARF locus with an indication of the primer pairs (PP) used in B–D. (B,C) ChIP assays using lysates from TIG3 BRAF-ER cells treated with or without OHT for the indicated time. In C, antibodies specific for JMJD3 or CBX8 were used; H3K27me3 was used in B. The degree of enrichment was measured by qPCR and results are presented as percentage of bound/input; the H3K27me3 signal was normalized to the levels of histone H3 bound. The error bars represent the standard deviation of the mean. qPCR primers amplifying last exon of the MTF2 gene were used as negative control. (D) Changes in K27me3 levels at the INK4A-TSS in TIG3 BRAF-ER cells treated with siJMJD3 or control in response to OHT treatment for 48 h. (E) Changes in H3K27me3 levels at the Ink4a-TSS in response to RasV12 expression with and without Jmjd3 knockdown in MEFs. (F) Array data from Basso et al. (2005) reanalyzed to show expression levels of JMJD3 in NB cells and different types of lymphoma and leukemia including DLBL, BL, HCL, primary effusion lymphoma (PEL), FL, MM, MCL, and B-CLL (BLL in the figure). The shaded area indicates the mean ± two standard deviations of controls (NB). Bars indicate the means; P-values of Mann-Whitney U-test are provided. (G) Model for the regulation of the INK4A–ARF locus by JMJD3. See the text for details.
References
-
- Agger K, Cloos PA, Christensen J, Pasini D, Rose S, Rappsilber J, Issaeva I, Canaani E, Salcini AE, Helin K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature. 2007;449:731–734. - PubMed
-
- Ameyar-Zazoua M, Wisniewska MB, Bakiri L, Wagner EF, Yaniv M, Weitzman JB. AP-1 dimers regulate transcription of the p14/p19ARF tumor suppressor gene. Oncogene. 2005;24:2298–2306. - PubMed
-
- Basso K, Margolin AA, Stolovitzky G, Klein U, Dalla-Favera R, Califano A. Reverse engineering of regulatory networks in human B cells. Nat Genet. 2005;37:382–390. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous