

Histone demethylase JMJD3 contributes to epigenetic control of INK4a/ARF by oncogenic RAS
- PMID: 19451218
- PMCID: PMC2685533
- DOI: 10.1101/gad.511109
Histone demethylase JMJD3 contributes to epigenetic control of INK4a/ARF by oncogenic RAS
Abstract
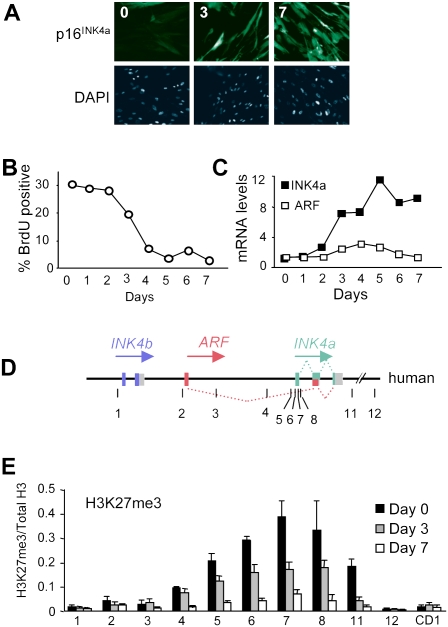
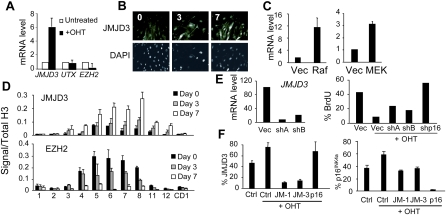
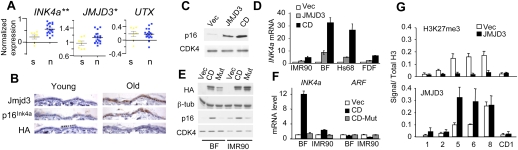
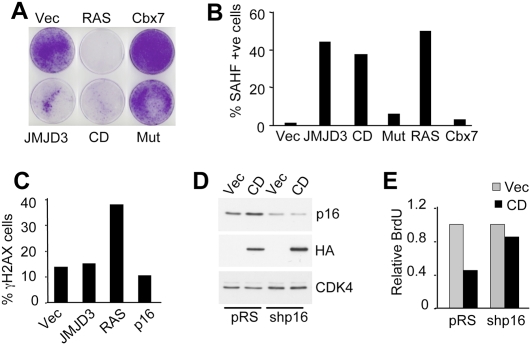
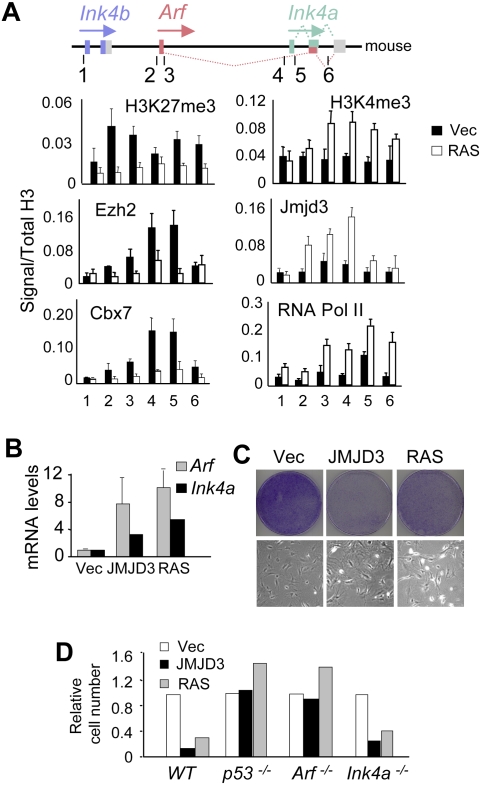
The INK4a/ARF tumor suppressor locus, a key executor of cellular senescence, is regulated by members of the Polycomb group (PcG) of transcriptional repressors. Here we show that signaling from oncogenic RAS overrides PcG-mediated repression of INK4a by activating the H3K27 demethylase JMJD3 and down-regulating the methyltransferase EZH2. In human fibroblasts, JMJD3 activates INK4a, but not ARF, and causes p16(INK4a)-dependent arrest. In mouse embryo fibroblasts, Jmjd3 activates both Ink4a and Arf and elicits a p53-dependent arrest, echoing the effects of RAS in this system. Our findings directly implicate JMJD3 in the regulation of INK4a/ARF during oncogene-induced senescence and suggest that JMJD3 has the capacity to act as a tumor suppressor.
Figures
References
-
- Acosta JC, O'Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, Fumagalli M, Da Costa M, Brown C, Popov N, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008;133:1006–1018. - PubMed
-
- Agger K, Cloos PA, Christensen J, Pasini D, Rose S, Rappsilber J, Issaeva I, Canaani E, Salcini AE, Helin K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature. 2007;449:731–734. - PubMed
-
- Ananthanarayanan M, Li S, Balasubramaniyan N, Suchy FJ, Walsh MJ. Ligand-dependent activation of the farnesoid X-receptor directs arginine methylation of histone H3 by CARM1. J Biol Chem. 2004;279:54348–54357. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous