

Myelin proteomics: molecular anatomy of an insulating sheath
- PMID: 19452287
- PMCID: PMC2758371
- DOI: 10.1007/s12035-009-8071-2
Myelin proteomics: molecular anatomy of an insulating sheath
Abstract
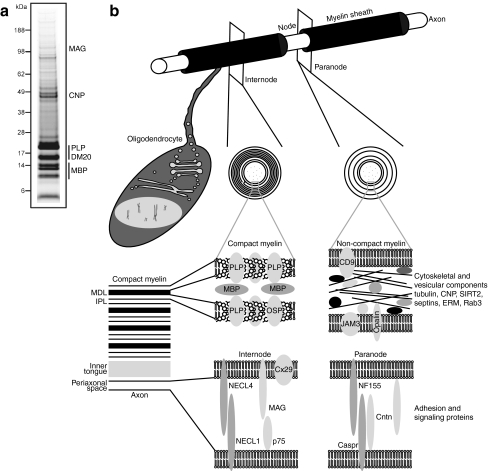
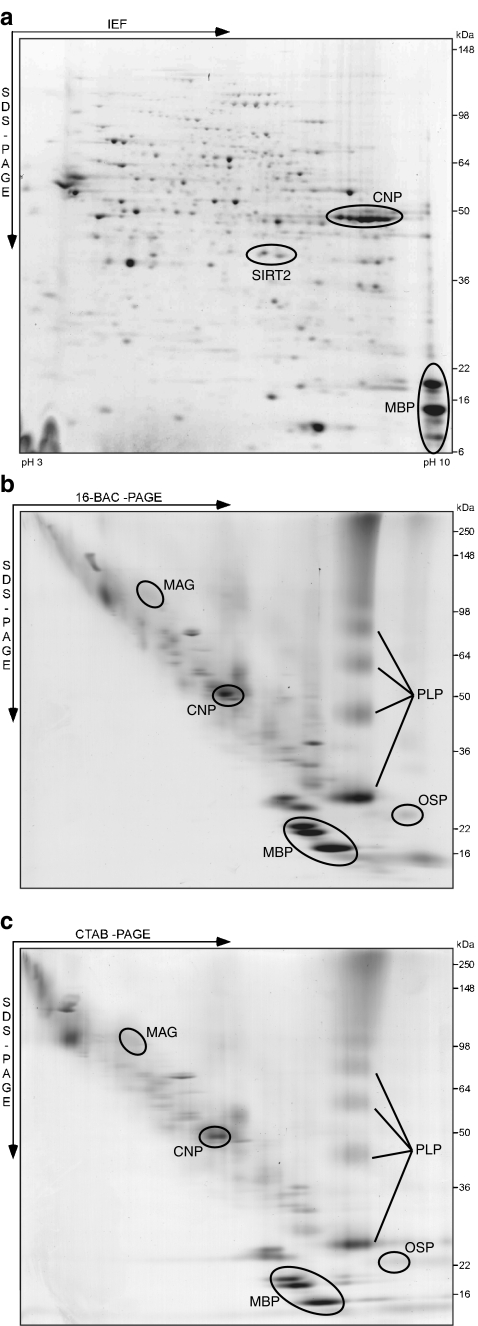
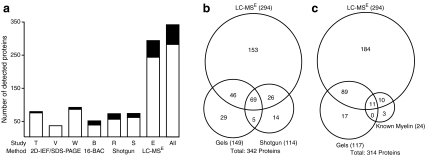
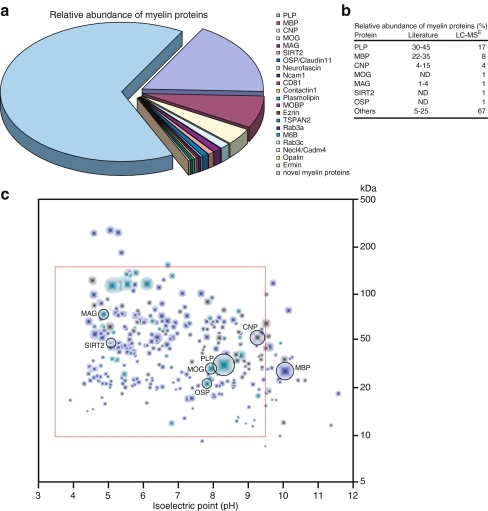
Fast-transmitting vertebrate axons are electrically insulated with multiple layers of nonconductive plasma membrane of glial cell origin, termed myelin. The myelin membrane is dominated by lipids, and its protein composition has historically been viewed to be of very low complexity. In this review, we discuss an updated reference compendium of 342 proteins associated with central nervous system myelin that represents a valuable resource for analyzing myelin biogenesis and white matter homeostasis. Cataloging the myelin proteome has been made possible by technical advances in the separation and mass spectrometric detection of proteins, also referred to as proteomics. This led to the identification of a large number of novel myelin-associated proteins, many of which represent low abundant components involved in catalytic activities, the cytoskeleton, vesicular trafficking, or cell adhesion. By mass spectrometry-based quantification, proteolipid protein and myelin basic protein constitute 17% and 8% of total myelin protein, respectively, suggesting that their abundance was previously overestimated. As the biochemical profile of myelin-associated proteins is highly reproducible, differential proteome analyses can be applied to material isolated from patients or animal models of myelin-related diseases such as multiple sclerosis and leukodystrophies.
Figures
References
-
- Hartline DK, Colman DR (2007) Rapid conduction and the evolution of giant axons and myelinated fibers. Curr Biol 17:R29–R35 - PubMed
-
- Poliak S, Peles E (2003) The local differentiation of myelinated axons at nodes of Ranvier. Nat Rev Neurosci 4:968–980 - PubMed
-
- Hildebrand C, Remahl S, Persson H, Bjartmar C (1993) Myelinated nerve fibres in the CNS. Prog Neurobiol 40:319–384 - PubMed
-
- Kirby BB, Takada N, Latimer AJ, Shin J, Carney TJ, Kelsh RN, Appel B (2006) In vivo time-lapse imaging shows dynamic oligodendrocyte progenitor behavior during zebrafish development. Nat Neurosci 9:1506–1511 - PubMed
-
- Brosamle C, Halpern ME (2002) Characterization of myelination in the developing zebrafish. Glia 39:47–57 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
