

Modulation of human immunodeficiency virus type 1 protease autoprocessing by charge properties of surface residue 69
- PMID: 19457992
- PMCID: PMC2708634
- DOI: 10.1128/JVI.00473-09
Modulation of human immunodeficiency virus type 1 protease autoprocessing by charge properties of surface residue 69
Abstract
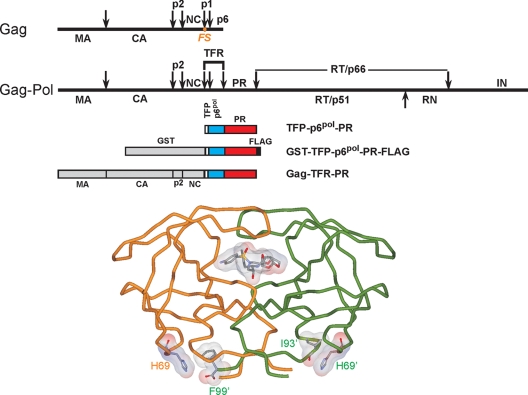
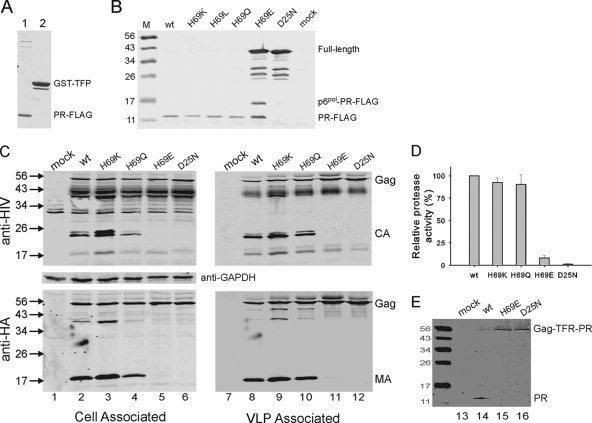
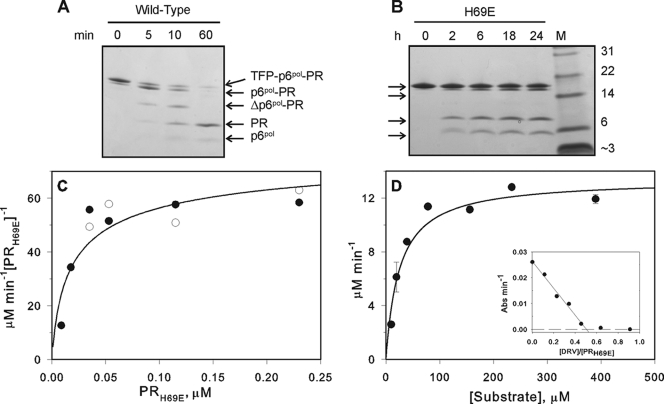
Mature, fully active human immunodeficiency virus protease (PR) is liberated from the Gag-Pol precursor via regulated autoprocessing. A chimeric protease precursor, glutathione S-transferase-transframe region (TFR)-PR-FLAG, also undergoes N-terminal autocatalytic maturation when it is expressed in Escherichia coli. Mutation of the surface residue H69 to glutamic acid, but not to several neutral or basic amino acids, impedes protease autoprocessing in bacteria and mammalian cells. Only a fraction of mature PR with an H69E mutation (PR(H69E)) folds into active enzymes, and it does so with an apparent Kd (dissociation constant) significantly higher than that of the wild-type protease, corroborating the marked retardation of the in vitro N-terminal autocatalytic processing of TFR-PR(H69E) and suggesting a folding defect in the precursor.
Figures
Similar articles
-
Flexible catalytic site conformations implicated in modulation of HIV-1 protease autoprocessing reactions.Retrovirology. 2011 Oct 10;8:79. doi: 10.1186/1742-4690-8-79. Retrovirology. 2011. PMID: 21985091 Free PMC article.
-
Cysteine 95 and other residues influence the regulatory effects of Histidine 69 mutations on Human Immunodeficiency Virus Type 1 protease autoprocessing.Retrovirology. 2010 Mar 23;7:24. doi: 10.1186/1742-4690-7-24. Retrovirology. 2010. PMID: 20331855 Free PMC article.
-
A Functional Interplay between Human Immunodeficiency Virus Type 1 Protease Residues 77 and 93 Involved in Differential Regulation of Precursor Autoprocessing and Mature Protease Activity.PLoS One. 2015 Apr 20;10(4):e0123561. doi: 10.1371/journal.pone.0123561. eCollection 2015. PLoS One. 2015. PMID: 25893662 Free PMC article.
-
Terminal interface conformations modulate dimer stability prior to amino terminal autoprocessing of HIV-1 protease.Biochemistry. 2012 Feb 7;51(5):1041-50. doi: 10.1021/bi201809s. Epub 2012 Jan 24. Biochemistry. 2012. PMID: 22242794 Free PMC article.
-
Human Immunodeficiency Virus Gag and protease: partners in resistance.Retrovirology. 2012 Aug 6;9:63. doi: 10.1186/1742-4690-9-63. Retrovirology. 2012. PMID: 22867298 Free PMC article. Review.
Cited by
-
Flexible catalytic site conformations implicated in modulation of HIV-1 protease autoprocessing reactions.Retrovirology. 2011 Oct 10;8:79. doi: 10.1186/1742-4690-8-79. Retrovirology. 2011. PMID: 21985091 Free PMC article.
-
Structural Studies of a Rationally Selected Multi-Drug Resistant HIV-1 Protease Reveal Synergistic Effect of Distal Mutations on Flap Dynamics.PLoS One. 2016 Dec 16;11(12):e0168616. doi: 10.1371/journal.pone.0168616. eCollection 2016. PLoS One. 2016. PMID: 27992544 Free PMC article.
-
High-affinity RNA Aptamers Against the HIV-1 Protease Inhibit Both In Vitro Protease Activity and Late Events of Viral Replication.Mol Ther Nucleic Acids. 2015 Feb 17;4(2):e228. doi: 10.1038/mtna.2015.1. Mol Ther Nucleic Acids. 2015. PMID: 25689224 Free PMC article.
-
Construction of a molecular clone of ovine enzootic nasal tumor virus.Virol J. 2016 Dec 30;13(1):209. doi: 10.1186/s12985-016-0660-x. Virol J. 2016. PMID: 28038674 Free PMC article.
-
Assay Development and Validation for Innovative Antiviral Development Targeting the N-Terminal Autoprocessing of SARS-CoV-2 Main Protease Precursors.Viruses. 2024 Jul 29;16(8):1218. doi: 10.3390/v16081218. Viruses. 2024. PMID: 39205192 Free PMC article.
References
-
- Barre-Sinoussi, F., J. C. Chermann, F. Rey, M. T. Nugeyre, S. Chamaret, J. Gruest, C. Dauguet, C. Axler-Blin, F. Vezinet-Brun, C. Rouzioux, W. Rozenbaum, and L. Montagnier. 1983. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 220868-871. - PubMed
-
- Cherry, E., C. Liang, L. Rong, Y. Quan, P. Inouye, X. Li, N. Morin, M. Kotler, and M. A. Wainberg. 1998. Characterization of human immunodeficiency virus type-1 (HIV-1) particles that express protease-reverse transcriptase fusion proteins. J. Mol. Biol. 28443-56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
