

Sparse but selective and potent synaptic transmission from the globus pallidus to the subthalamic nucleus
- PMID: 19458148
- PMCID: PMC2712268
- DOI: 10.1152/jn.00305.2009
Sparse but selective and potent synaptic transmission from the globus pallidus to the subthalamic nucleus
Abstract
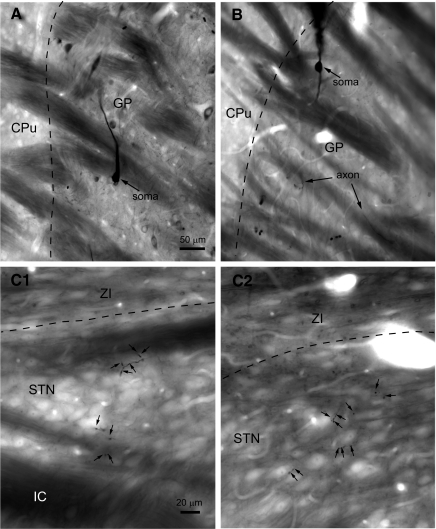
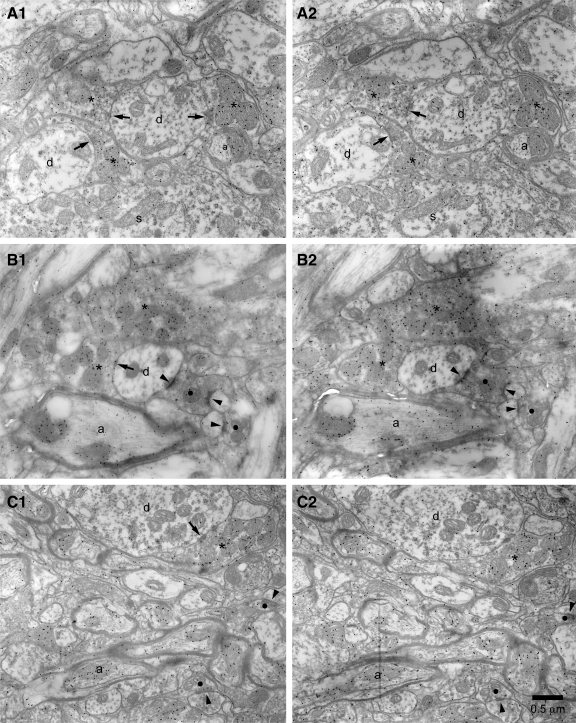
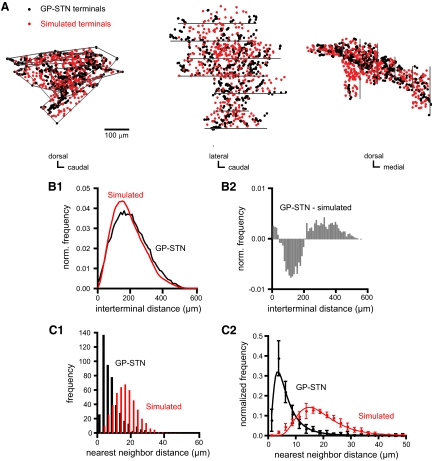
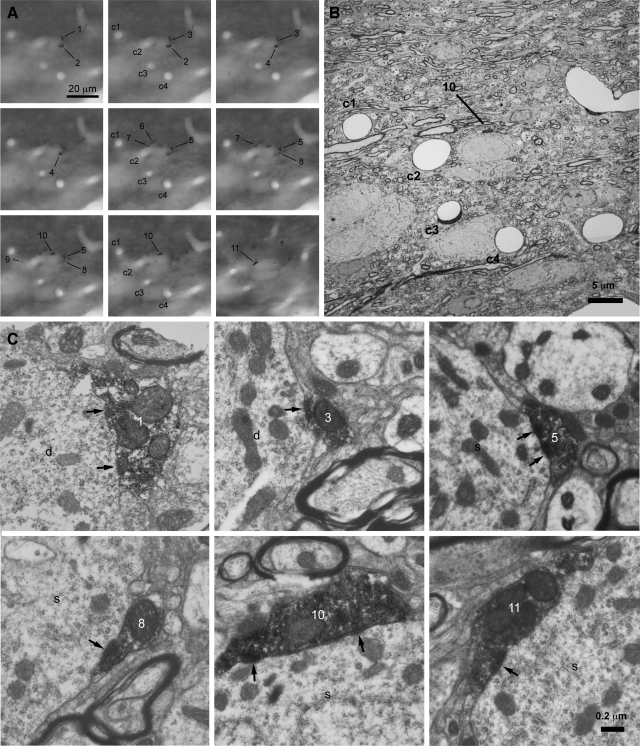
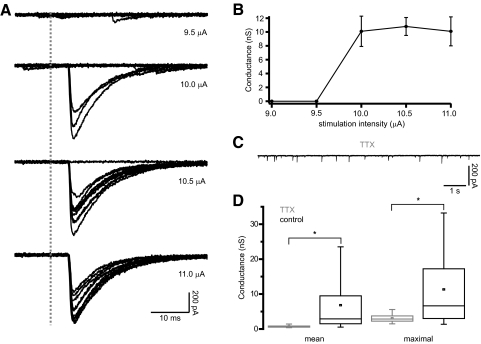
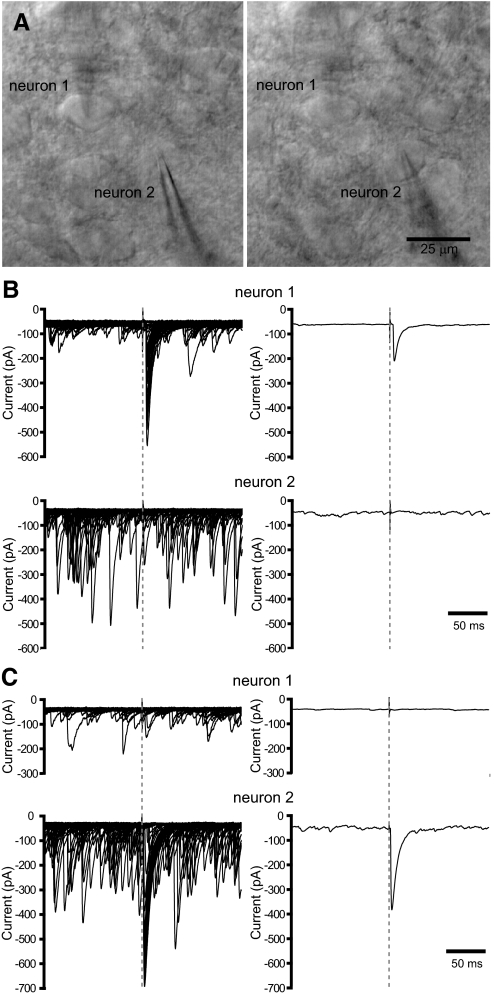
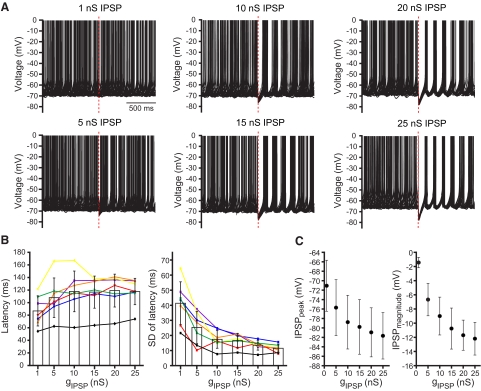
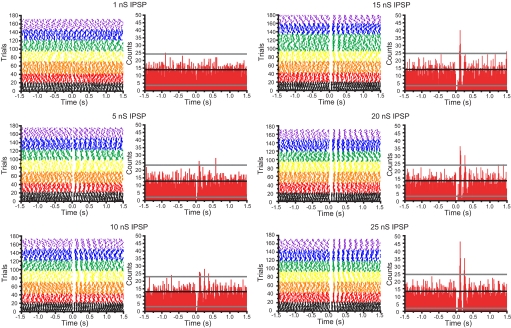
The reciprocally connected GABAergic globus pallidus (GP)-glutamatergic subthalamic nucleus (STN) network is critical for voluntary movement and an important site of dysfunction in movement disorders such as Parkinson's disease. Although the GP is a key determinant of STN activity, correlated GP-STN activity is rare under normal conditions. Here we define fundamental features of the GP-STN connection that contribute to poorly correlated GP-STN activity. Juxtacellular labeling of single GP neurons in vivo and stereological estimation of the total number of GABAergic GP-STN synapses suggest that the GP-STN connection is surprisingly sparse: single GP neurons maximally contact only 2% of STN neurons and single STN neurons maximally receive input from 2% of GP neurons. However, GP-STN connectivity may be considerably more selective than even these estimates imply. Light and electron microscopic analyses revealed that single GP axons give rise to sparsely distributed terminal clusters, many of which correspond to multiple synapses with individual STN neurons. Application of the minimal stimulation technique in brain slices confirmed that STN neurons receive multisynaptic unitary inputs and that these inputs largely arise from different sets of GABAergic axons. Finally, the dynamic-clamp technique was applied to quantify the impact of GP-STN inputs on STN activity. Small fractions of GP-STN input were sufficiently powerful to inhibit and synchronize the autonomous activity of STN neurons. Together these data are consistent with the conclusion that the rarity of correlated GP-STN activity in vivo is due to the sparsity and selectivity, rather than the potency, of GP-STN synaptic connections.
Figures
Similar articles
-
Globus pallidus neurons dynamically regulate the activity pattern of subthalamic nucleus neurons through the frequency-dependent activation of postsynaptic GABAA and GABAB receptors.J Neurosci. 2005 Jul 6;25(27):6304-15. doi: 10.1523/JNEUROSCI.0450-05.2005. J Neurosci. 2005. PMID: 16000620 Free PMC article.
-
D2-like dopamine receptor-mediated modulation of activity-dependent plasticity at GABAergic synapses in the subthalamic nucleus.J Physiol. 2008 Apr 15;586(8):2121-42. doi: 10.1113/jphysiol.2008.151118. Epub 2008 Feb 21. J Physiol. 2008. PMID: 18292127 Free PMC article.
-
Autonomous initiation and propagation of action potentials in neurons of the subthalamic nucleus.J Physiol. 2008 Dec 1;586(23):5679-700. doi: 10.1113/jphysiol.2008.155861. Epub 2008 Oct 2. J Physiol. 2008. PMID: 18832425 Free PMC article.
-
GABAergic control of the subthalamic nucleus.Prog Brain Res. 2007;160:173-88. doi: 10.1016/S0079-6123(06)60010-1. Prog Brain Res. 2007. PMID: 17499114 Review.
-
Globus pallidus external segment.Prog Brain Res. 2007;160:111-33. doi: 10.1016/S0079-6123(06)60007-1. Prog Brain Res. 2007. PMID: 17499111 Review.
Cited by
-
Coordinated reset stimulation in a large-scale model of the STN-GPe circuit.Front Comput Neurosci. 2014 Nov 27;8:154. doi: 10.3389/fncom.2014.00154. eCollection 2014. Front Comput Neurosci. 2014. PMID: 25505882 Free PMC article.
-
Dopamine depletion leads to pathological synchronization of distinct basal ganglia loops in the beta band.PLoS Comput Biol. 2023 Apr 27;19(4):e1010645. doi: 10.1371/journal.pcbi.1010645. eCollection 2023 Apr. PLoS Comput Biol. 2023. PMID: 37104542 Free PMC article.
-
Connectivity and Functionality of the Globus Pallidus Externa Under Normal Conditions and Parkinson's Disease.Front Neural Circuits. 2021 Mar 2;15:645287. doi: 10.3389/fncir.2021.645287. eCollection 2021. Front Neural Circuits. 2021. PMID: 33737869 Free PMC article.
-
Short-term depression of external globus pallidus-subthalamic nucleus synaptic transmission and implications for patterning subthalamic activity.J Neurosci. 2013 Apr 24;33(17):7130-44. doi: 10.1523/JNEUROSCI.3576-12.2013. J Neurosci. 2013. PMID: 23616523 Free PMC article.
-
Proliferation of external globus pallidus-subthalamic nucleus synapses following degeneration of midbrain dopamine neurons.J Neurosci. 2012 Oct 3;32(40):13718-28. doi: 10.1523/JNEUROSCI.5750-11.2012. J Neurosci. 2012. PMID: 23035084 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources