

Parainfluenza virus 5 genomes are located in viral cytoplasmic bodies whilst the virus dismantles the interferon-induced antiviral state of cells
- PMID: 19458173
- PMCID: PMC2885057
- DOI: 10.1099/vir.0.012047-0
Parainfluenza virus 5 genomes are located in viral cytoplasmic bodies whilst the virus dismantles the interferon-induced antiviral state of cells
Abstract
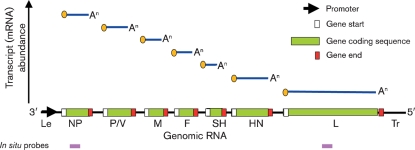
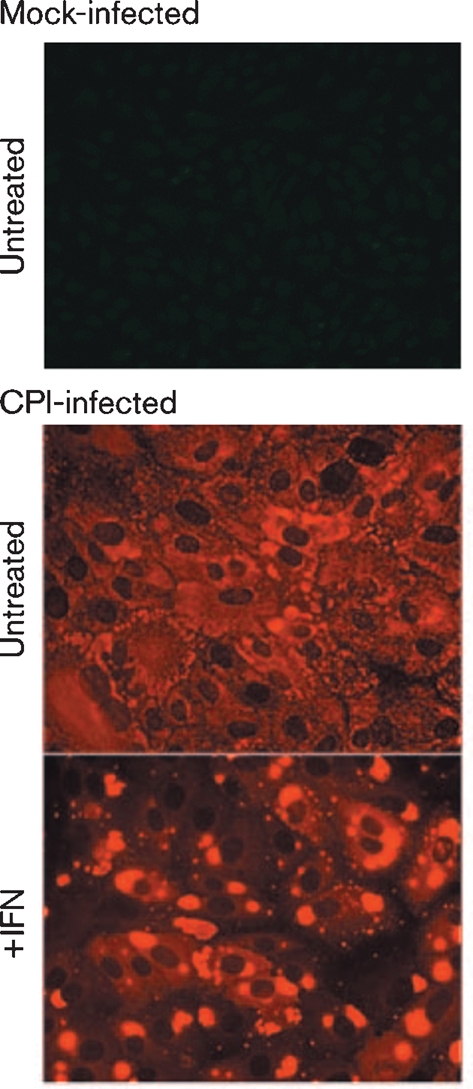
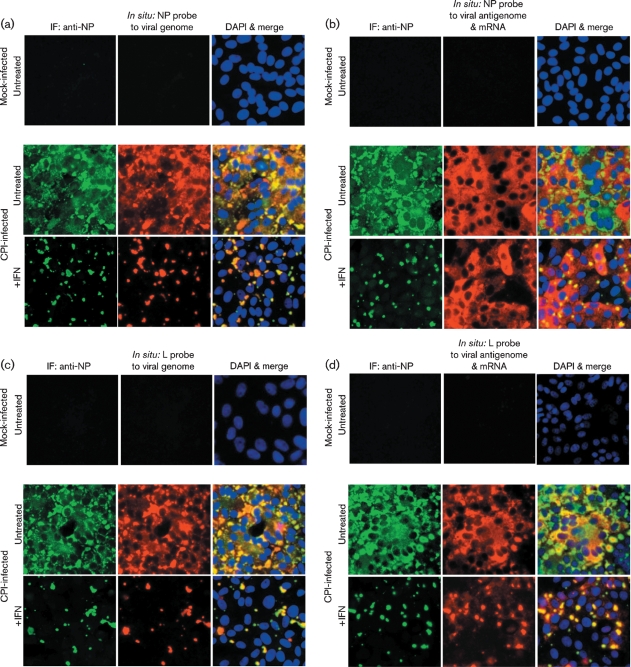
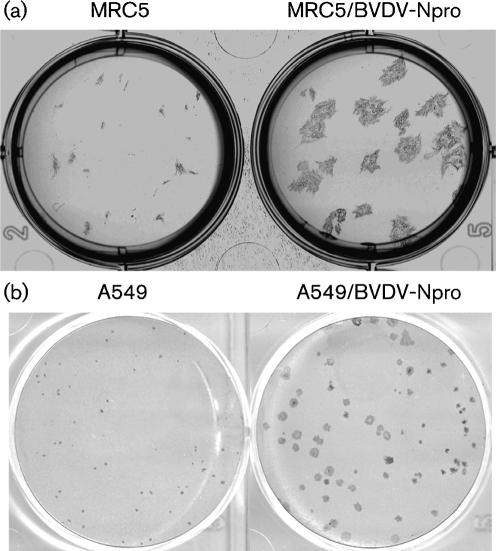
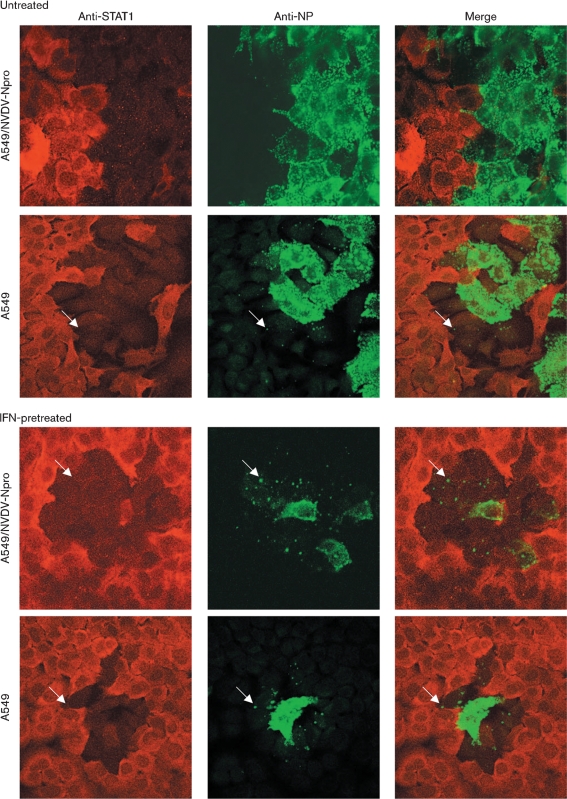
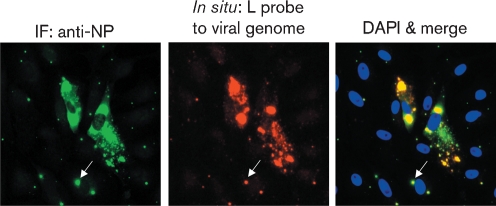
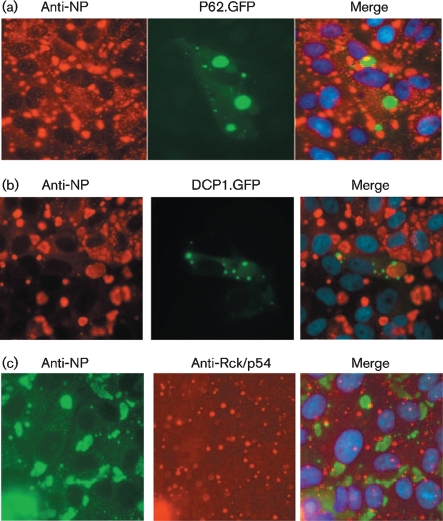
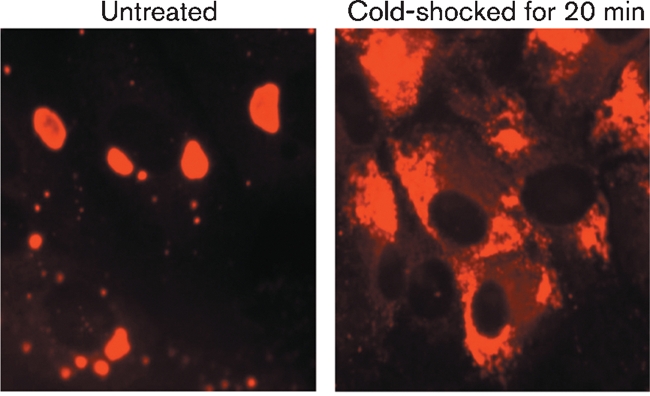
Although the replication cycle of parainfluenza virus type 5 (PIV5) is initially severely impaired in cells in an interferon (IFN)-induced antiviral state, the virus still targets STAT1 for degradation. As a consequence, the cells can no longer respond to IFN and after 24-48 h, they go out of the antiviral state and normal virus replication is established. Following infection of cells in an IFN-induced antiviral state, viral nucleocapsid proteins are initially localized within small cytoplasmic bodies, and appearance of these cytoplasmic bodies correlates with the loss of STAT1 from infected cells. In situ hybridization, using probes specific for the NP and L genes, demonstrated the presence of virus genomes within these cytoplasmic bodies. These viral cytoplasmic bodies do not co-localize with cellular markers for stress granules, cytoplasmic P-bodies or autophagosomes. Furthermore, they are not large insoluble aggregates of viral proteins and/or nucleocapsids, as they can simply and easily be dispersed by 'cold-shocking' live cells, a process that disrupts the cytoskeleton. Given that during in vivo infections, PIV5 will inevitably infect cells in an IFN-induced antiviral state, we suggest that these cytoplasmic bodies are areas in which PIV5 genomes reside whilst the virus dismantles the antiviral state of the cells. Consequently, viral cytoplasmic bodies may play an important part in the strategy that PIV5 uses to circumvent the IFN system.
Figures
References
-
- Baumgartner, W., Krakowka, S. & Blakeslee, J. R. (1987). Persistent infection of Vero cells by paramyxoviruses. A morphological and immunoelectron microscopic investigation. Intervirology 27, 218–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous