

Gap junction turnover is achieved by the internalization of small endocytic double-membrane vesicles
- PMID: 19458184
- PMCID: PMC2710821
- DOI: 10.1091/mbc.e09-04-0288
Gap junction turnover is achieved by the internalization of small endocytic double-membrane vesicles
Abstract
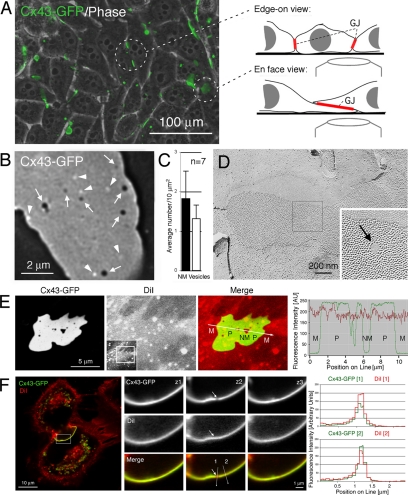
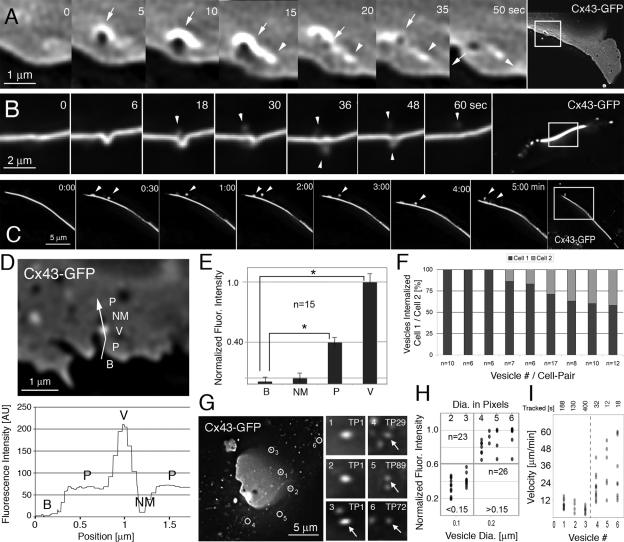
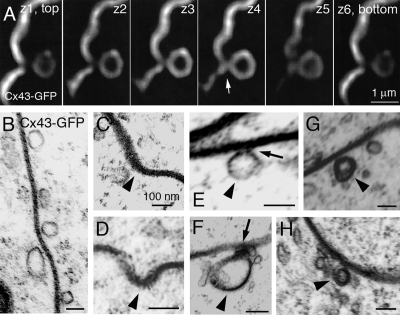
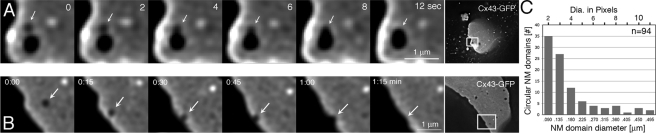
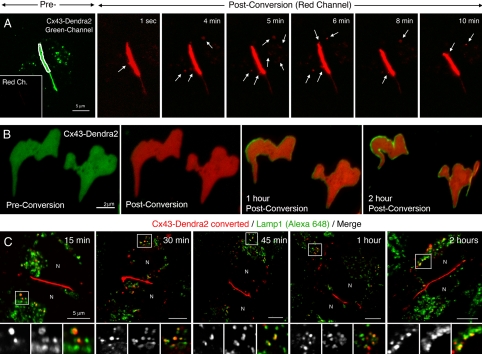
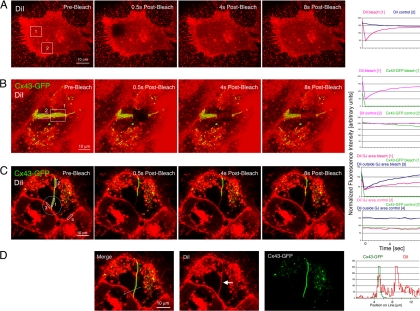
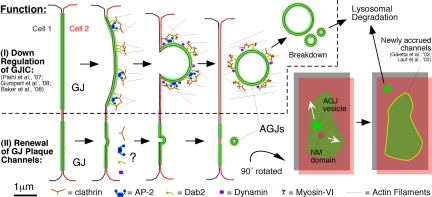
Double-membrane-spanning gap junction (GJ) channels cluster into two-dimensional arrays, termed plaques, to provide direct cell-to-cell communication. GJ plaques often contain circular, channel-free domains ( approximately 0.05-0.5 mum in diameter) identified >30 y ago and termed nonjunctional membrane (NM) domains. We show, by expressing the GJ protein connexin43 (Cx43) tagged with green fluorescent protein, or the novel photoconvertible fluorescent protein Dendra2, that NM domains appear to be remnants generated by the internalization of small GJ channel clusters that bud over time from central plaque areas. Channel clusters internalized within seconds forming endocytic double-membrane GJ vesicles ( approximately 0.18-0.27 mum in diameter) that were degraded by lysosomal pathways. Surprisingly, NM domains were not repopulated by surrounding channels and instead remained mobile, fused with each other, and were expelled at plaque edges. Quantification of internalized, photoconverted Cx43-Dendra2 vesicles indicated a GJ half-life of 2.6 h that falls within the estimated half-life of 1-5 h reported for GJs. Together with previous publications that revealed continuous accrual of newly synthesized channels along plaque edges and simultaneous removal of channels from plaque centers, our data suggest how the known dynamic channel replenishment of functional GJ plaques can be achieved. Our observations may have implications for the process of endocytic vesicle budding in general.
Figures
References
-
- Beardslee M. A., Laing J. G., Beyer E. C., Saffitz J. E. Rapid turnover of connexin43 in the adult rat heart. Circ. Res. 1998;83:629–635. - PubMed
-
- Berthoud V. M., Minogue P. J., Laing J. G., Beyer E. C. Pathways for degradation of connexins and gap junctions. Cardiovasc. Res. 2004;62:256–267. - PubMed
-
- Bruzzone R., White T. W., Paul D. L. Connections with connexins: the molecular basis of direct intercellular signaling. Eur. J. Biochem. 1996;238:1–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
